Правила обрезки и формирования для хвойных деревьев и кустарников
Ежегодная обрезка хвойных растений, если они здоровы, не требуется. Обрезку применяют в тех случаях, когда необходимо удалить поврежденные и засохшие ветви, или же при желании придать растению определенную форму.
Хвойные хорошо стригутся. Но следует понимать, что это все таки стресс для растения, поэтому не рекомендуется уменьшать объем более чем на 1/3. Обычно вполне достаточно подчеркнуть природную форму растения.
Для формирования более плотной густой кроны, обрезают около 1/3 годового прироста. Следует учитывать, так как хвойники не имеют спящих почек, то голые ветви без хвои обрезают, иначе они засохнут. Через год после посадки/пересадки, не ранее, можно приступать к формированию растения.
Обрезку производят только острым и чистым инструментом (секатор, ножовка и т.п.), чтобы не занести инфекцию. Затем срезы желательно обработать лаком, марганцовкой или садовым варом.
Проводить стрижку можно с весны до осени, но наиболее подходящим считается период с конца февраля по март, до начала сокодвижения. Но каждый вид растения индивидуален, и имеет свои периоды динамичного роста, в это время обильно выделяется смола, и стрижка не желательна.
Но каждый вид растения индивидуален, и имеет свои периоды динамичного роста, в это время обильно выделяется смола, и стрижка не желательна.
Такие растения, как кипарисовик, кипарис, лох узколистный, лавровишня, некоторые виды можжевельника практически не подрезают. Им рекомендована слабая обрезка, для укорачивания некоторых ветвей, которые слишком неряшливо выступают из кроны.
А, например, туя западная, тис ягодный, обыкновенная ель, кипарисовик Лавсона и другие виды можжевельников наоборот хорошо переносят стрижку. Обычно их стригут не более 2-х раз в год. Обрезают более длинную часть ветви, для того чтобы боковая заняла ее место. Начинают стричь растение ранней весной (март-апрель), до начала роста побегов, потом еще раз осенью (июль-август), для того, чтобы новые побеги до зимы успели одревеснеть. Желательно стрижку проводить в пасмурные дни. Хвойники этой группы, для формирования густой плотной кроны, обрезают на 2-й год после посадки, удаляя 2/3 длины боковых побегов и 1/3 верхушечного побега. Каждый раз поднимая верхушку до нужной высоты. Такая стрижка способствует образованию боковых ветвей, что создаст густую зеленую изгородь.
Каждый раз поднимая верхушку до нужной высоты. Такая стрижка способствует образованию боковых ветвей, что создаст густую зеленую изгородь.
Так как можжевельники медленнорастущие, то обрезать их рекомендуют по необходимости, удаляя поврежденные, сухие ветви и укорачивая слишком длинные, выступающие, создающие сильную асимметрию. Период стрижки можжевельников — начало мая или начало октября (период покоя растения).
В общем, можжевельники неплохо переносят формировку. Кустовидные многоствольные сорта подойдут для создания стриженной живой изгороди, а раскидистые, стелющиеся виды можно укорачивать для формирования более компактной формы.
Другие хвойные, такие как пихта, ель, сосна, обрезаются после появления новых приростов, ровно на половину. Если необходимо поддержать существующий размер хвойного растения, то молодые побеги удаляют.
Отдельно стоит выделить сосну. Дело в том, что у сосны побеги формируются только из почек, расположенных на концах ветвей, весной (в исключительных случаях на самой древесине). Поэтому обрезая какую-либо одревесневшую ветвь, она уже ветвиться не будет, останется просто сук.
Поэтому обрезая какую-либо одревесневшую ветвь, она уже ветвиться не будет, останется просто сук.
Если сосна уже подросла, то ее формируют, укорачивая слишком длинные ветви, для роста боковых заменяющих ветвей, так, чтобы не оставлять сучков. Верхушечный побег можно укоротить до разветвления ветвей, а в последствии, не давать возможности формирования лидирующего побега.
Сосну нужно начинать формировать с самого начала, так как в начале роста она имеет компактную аккуратную форму, прибавляя по 15 см, затем после образования сильного стержневого корня она начинает быстро расти вверх, сильно вытягиваясь.
Для замедления роста и придания более декоративного, компактного вида и более густой кроны, сосну необходимо прищипывать (укорачивая молодые, еще не одревесневшие побеги). Так как молодые приросты растут с мая до середины июня, примерно в это время прищипку делают на самом начале стадии отхода иголочек от молодого побега, в момент остановки его роста. Для сосны обыкновенной начало формировки (прищипки) приходится на достижение ею 1-1,5-метровой высоты.
описание пихты с синими шишками, посадка и уход, виды и сорта, выращивание. Как быстро растет?
В среде садоводов корейская пихта пользуется заслуженной популярностью благодаря своей неприхотливости, зимостойкости, выносливости и декоративному виду, сохраняющемуся круглый год. Дерево активно используется в ландшафтном дизайне, кроме того, высоко оценены его лечебные качества, способствующие обеззараживанию воздуха и устранению инфекций и вредителей на участке.
Описание
Пихта Abies koreana – дерево, относящееся к семейству сосновых. В природной среде произрастает в северных регионах, хвойных и смешанных лесных массивах, на склонах гор, достигая высоты 15 м и более.

- Крона имеет конусовидную, иногда пирамидальную форму, а окружность её ствола составляет 0,5-0,8 м.
- Отличительная особенность хвойника – внушительные размеры кроны, культура быстро растёт вширь, но отличается медленным ростом в высоту.
- Вечнозелёная пихта корейская – обладательница хорошо развитой и мощной корневой системы.
- Кора у небольших пихт серо-серебристая, тонкая и гладкая, но с возрастом наращивает толщину, приобретает красновато-коричневый оттенок, и покрывается трещинами.
- Молодые побеги имеют желтоватый цвет, но с годами он меняется на фиолетовый. Формирующиеся на них почки имеют шарообразную форму и содержат большое количество пахучей смолы с приятным хвойным ароматом.
- Цвет хвои яркий, изумрудный на внешней стороне иголок, а на внутренней стороне иглы имеют серебристые прожилки.
- Края хвоинок слегка загнуты – на молодых деревцах они колючие и жёсткие, у взрослых экземпляров обладают более сглаженным, тупым концом.
 Длина игл – 20 мм, а ширина – около 2,5 мм.
Длина игл – 20 мм, а ширина – около 2,5 мм. - Красиво выглядят ветки пихты с синими шишками, имеющими пурпурный оттенок и цилиндрическую форму. Плоды дерева довольно крупные – до7-8 см в длину и до 3 см в диаметре. Обычно формирование их происходит в верхней части кроны на свежих побегах.
- Когда семена созревают, закрывающие их чешуйки твердеют, проходя процесс одревеснения, и опадают, а семенной материал разносится ветром. Кстати, опыление шишек тоже происходит за счёт движения воздушных масс.
 При неблагоприятных условиях и неудовлетворительном уходе рост её затормаживается и она не достигает оптимальной высоты. В результате растение больше похоже на кустарник, имеющий округлую форму кроны.
При неблагоприятных условиях и неудовлетворительном уходе рост её затормаживается и она не достигает оптимальной высоты. В результате растение больше похоже на кустарник, имеющий округлую форму кроны.Культура хорошо переносит минусовые температуры, но некомфортно чувствует себя при засушливой и жаркой погоде. В отличие от некоторых хвойных пород данный вид чувствителен к любым изменением в составе воздуха, особенно к загазованности, типичной для города. Это исключает возможность её посадки вдоль шоссе и автомобильных трасс.
Этот хвойный вид считается полезной культурой, а садоводы ценят его не только за красоту некоторых разновидностей, но и за способность дерева очищать воздух и предотвращать нападение вредных насекомых благодаря летучим дезинфицирующим веществам – фитонцидам, выделяемым хвоей.
Виды и сорта
Растение имеет большое количество искусственно выведенных сортов и видов, которые подходят для озеленения и оформления как частных территорий, так и городских парков, скверов.
Ниже перечислены наиболее востребованные культуры, подходящие для самых разных регионов.
- Миниатюрное деревце «Компакта» относится к карликовой разновидности корейской пихты и способно вырасти до 80 см, прибавляя в год по 5-6 см. У неё длинная красивая хвоя серебристо-синего цвета, являющаяся ее основной декоративной ценностью и компенсирующей редкое формирование шишек.
Хвойник можно выращивать в вазонах, использовать для украшения веранд и парадного входа. Также посадками пихт обрамляют садовые дорожки, высаживают в составе композиций, включающих искусственные пруды и камни.
- Карликовый кустарник «Тундра» имеет окружность кроны 0,6 м и высоту 0,4 м. Хорошо приспособлен к выращиванию в тени и любит увлажнённую почву.
 Отличается мягкой, короткой и блестящей хвоей яркого зелёного цвета с серебристой нижней стороной. Сорт применяется для оформления вересковых и каменистых садов, альпинариев, озеленения склонов и крыш.
Отличается мягкой, короткой и блестящей хвоей яркого зелёного цвета с серебристой нижней стороной. Сорт применяется для оформления вересковых и каменистых садов, альпинариев, озеленения склонов и крыш.
- «Сильвер Стар» – вечнозелёное растение с формой кроны в виде неправильного конуса. Пихта способна вырасти до 1 м при окружности зелёной части 70-80 см. В год вырастает до 8 см. Хвоинки ярко-изумрудные снаружи, внутренняя их сторона имеет белый или серебристый цвет. Компактное дерево уже в молодом возрасте выбрасывает красивые фиолетовые шишки. Использование предусматривает одиночные посадки и композиции с цветами, хвойными деревьями, травянистыми растениями.
- Голубой карлик «Блю Эмперор» отличается подушковидной неправильной формой кроны.
 Хвоинки серо-голубые, закруглённые на концах. В высоту сорт достигает не больше 1,5 м, а в год растёт на 6-8 см. Нередко крона его стелется по земле, но побеги растут вертикально вверх. Культура зимостойкая (зона 5а), нуждается в рыхлой плодородной почве. Вид Blue Emperor нашёл своё применение при создании альпийских горок, оформлении каменистых участков.
Хвоинки серо-голубые, закруглённые на концах. В высоту сорт достигает не больше 1,5 м, а в год растёт на 6-8 см. Нередко крона его стелется по земле, но побеги растут вертикально вверх. Культура зимостойкая (зона 5а), нуждается в рыхлой плодородной почве. Вид Blue Emperor нашёл своё применение при создании альпийских горок, оформлении каменистых участков.
- «Блю Мэджик» – небольшое медленно растущее дерево, которое во взрослом состоянии имеет высоту около 1 м, но с годами способно вырасти до 2,5 м. Форма кроны может быть конусообразной и подушковидной. Особенность сорта – зелёно-голубой цвет хвои при теневом выращивании. При хорошем освещении пихта имеет серебристо-голубой окрас. Высаживается отдельно, как элемент садовых композиций или в групповых посадках.
- «Оберон» – этот сорт пихты имеет редкие декоративные качества. Его хвоя имеет широкие и короткие иглы, ярко-зелёные снаружи, и белые снизу. Растение по мере роста достигает одинаковой ширины и высоты – 1,5 м, и напоминает конус неправильной формы. Деревце подходит для озеленения альпийских горок и небольших садовых ансамблей.
- Разновидность корейской пихты «Ауреа» тоже считается карликовой. По мере роста она достигает 1,5 метра, а окончательная её высота – около 4-4,5 м. Сорт славится волшебным золотистым окрасом ветвей в весеннее время, на молодых ветках растения уже могут образовываться красивые шишки пурпурного цвета.
 Крону дерева можно формировать при помощи обрезки. По зимостойкости вид относится к зоне 4, но нуждается в защите от ветра.
Крону дерева можно формировать при помощи обрезки. По зимостойкости вид относится к зоне 4, но нуждается в защите от ветра.
- Сорт «Дискус» – один из самых медленнорастущих видов, его прирост в год составляет всего 3 см. Культуру отличает бело-зелёная крона конусовидной формы. Взрослые экземпляры вытягиваются до 1,2 м в высоту и достигают 90 см в ширину. Пихта чувствительна к сквознякам, но не боится сильных морозов.
В качестве декоративного украшения используется в создании рокариев, альпийских горок, допускается выращивание культуры в контейнерах.
- Пихта «Инга» поражает своим цветом, близким к оттенку морской волны. Его яркость смягчена характерными серебристыми полосками. Крона в виде конуса пышная и густая, но за счёт отсутствия чёткой геометрии смотрится особенно оригинально.
 Деревце медленно растёт, но будучи взрослым, вырастает до 0,8-1 м, хотя к 30 годам может достигнуть 3-4 м. Применяется сорт для посадки в группе, для создания парковых и садовых ансамблей, живых изгородей.
Деревце медленно растёт, но будучи взрослым, вырастает до 0,8-1 м, хотя к 30 годам может достигнуть 3-4 м. Применяется сорт для посадки в группе, для создания парковых и садовых ансамблей, живых изгородей.
- Корейский сорт «Цис» имеет округлую или шаровидную крону, мягкую изумрудно-зелёную хвою. Его пурпурно-лиловые шишки длиной в 5 см растут вверх. Хвойный карлик каждый год вырастает на 1-3 см и имеет диаметр кроны в 1 м. Этот вид лучше всего переносит засушливую погоду, но требует хорошего дренажа почвы. Зелёный «гном» станет идеальным декоративным элементом в любой садовой композиции. Сорт хорошо выглядит при групповой посадке и в сочетании с другими хвойными и лиственными растениями, включая цветы и травы.
- Пихта «Таня» – красивый сорт, который обладает хвоей светло-зелёного цвета с серебристым оттенком.
 Несмотря на небольшую высоту (1 м), миниатюрный вид может похвастаться большими шишками лилового цвета с пурпурным отливом.
Несмотря на небольшую высоту (1 м), миниатюрный вид может похвастаться большими шишками лилового цвета с пурпурным отливом.
Крона дерева поддаётся художественной обрезке, её можно формировать по своему желанию и в соответствии с задуманным дизайном участка. Хвойник отличает морозостойкость, способность расти на не слишком плодородных грунтах. Сорт применяют в комбинации с низкорослыми хвойниками и лиственными кустарниками.
Для различных видов корейской пихты характерны небольшие размеры, хотя некоторые разновидности в 30-40 лет могут вырастать на приличную высоту.
Но поскольку деревья растут медленно, это непременные участники садовых ансамблей и отлично справляются с ролью компактных декоративных украшений параллельно с другими растениями.

Посадка
При выращивании культуры необходимо определиться с подходящим местом для её посадки. Несмотря на неприхотливость растения, сильно затенённое место для него не подходит, к тому же пихта может потерять свой красивый декоративный цвет хвои. Поэтому желательно высаживать хвойник на достаточно открытом месте, а затем искусственно притенять молодые саженцы в первое время.
Хорошо, если пихта будет высажена на просторном участке, но защищена от ветров стоящими рядом постройками. При разном использовании дерева надо учитывать некоторые нюансы его размещения:
- если корейская пихта послужит в качестве одного из декоративных компонентов рокария или альпинария, между саженцами соблюдается интервал 3 м;
- при создании аллеи расстояние между деревьями должно составлять не меньше 4 м;
- когда культура предназначена для использования в живой изгороди, между растениями оставляют 2,5 м.
Суглинистая, супесчаная почва с низкой кислотностью больше всего подходит для этого растения. За две недели до посадки её перекапывают, а если необходимо обогатить грунт, добавляют в него плодородной садовой земли и торфа, при повышенной кислотности проводят известкование. Тяжёлую землю, состоящую из глины, разбавляют песком. В небольшом количестве вносят органические удобрения (компост, перегной, куриный помёт – около 8 г на 1 кв. м). Можно подкормить грунт препаратами калия и фосфора, а также мочевины (20 г на 1 кв. м).
Саженцы для высадки приобретают в возрасте 1 года, обязательно с закрытой корневой системой, которая может адаптироваться в открытом грунте. Такие молодые пихты продаются в пакетах, мешковине или контейнерах.
Непосредственно перед размещением в земле корни погружают на 2 часа в раствор биостимулятора – «Корневина», «Эпина» или «Фитоспорина».
Растения лучше сажать в апреле, хотя допускается и осенняя посадка. Правильно делать это при наступлении устойчивой тёплой погоды и отсутствии ночных заморозков.
Последовательность действий:
- посадочная лунка выкапывается диаметром в 60 см и с такой же глубиной;
- дренажный слой можно соорудить из кусочков кирпича, керамзита, гальки и песка;
- почвосмесь готовят из торфа, дерновой земли, песка и перегноя, взятых в одинаковом количестве, наполняют ямку на 2/3, добавляя «Нитроаммофоску» (300 г), после этого обильно поливают отстоянной водой;
- растение размещают по центру лунки, расправив его корневые отростки; корневую шейку оставляют над поверхностью земли;
- плодородный грунт подсыпают постепенно и одновременно уплотняют его, после чего поливают тёплой талой водой.
В конце проводят мульчирование перегноем, древесной щепой и опилками, слой которых составляет не менее 8 см.

Уход
Корейский хвойник, разумеется, может выживать и в неблагоприятных условиях, но для того, чтобы сохранить его здоровье и декоративный вид, за ним надо ухаживать.
Забота о культуре предусматривает различные аграрные работы.
- Увлажнённая почва необходима для молодых растений, поэтому первые 2-3 года растения поливают регулярно, не допуская пересушивания приствольной земли (около 10 л на 1 саженец). В засушливый период взрослым пихтам требуется до 20 л воды еженедельно. Также деревца следует опрыскивать рано утром и после заката солнца, чтобы не вызвать ожогов на хвое.
- Два года подряд после посадки почву под молодыми пихтами регулярно пропалывают от сорняков, рыхлят при помощи тяпки не глубже 6-7 см в глубину, присыпают мульчей, предупреждающей пересыхание, застой воды и развитие заболеваний.

- Во многом успешное выращивание саженцев зависит от состава грунта. Во время высаживания они уже получают порцию полезных веществ, после этого внесение удобрений понадобится на будущий год после таяния снегов и с наступлением вегетации. Для этого используют такую агропродукцию, как «Кемира». Во время плодоношения, то есть появления шишек, растение также нужно подкормить, желательно минеральными препаратами для хвойников («Гера», «Мягкая сила», «Бона Форте»).
У декоративных и карликовых видов корейской пихты нет необходимости формировать крону, однако они нуждаются в весенней и осенней санитарной обрезке, когда удаляются больные, засохшие и повреждённые веточки. Весной процедуру проводят до начала сокодвижения. Исключение составляет обрезание макушки некоторых деревьев, чтобы усилить ветвление.
На зиму для молодых пихт создают укрытие из нетканых материалов, предварительно возведя деревянный каркас, к которому их фиксируют. В приствольную зону насыпают торф, перегной, листовой опад и опилки.
Размножение
Разводить различные сорта пихты можно при помощи трёх способов. Один из них – выращивание из семян. Сделать это довольно сложно, потому что трудно собрать разлетающиеся лёгкие семена. Поэтому берут ещё не созревшие шишки и высушивают их. После этого извлекают семена и содержат их в прохладе при повышенной влажности.
Сажать семена можно в грунт или в контейнеры с тем расчетом, чтобы на следующий год поместить в открытую почву. Закалённые в холоде семечки высаживают в землю в апреле на глубину до 2 см, используя питательный субстрат для хвойников. Рассаде создают подходящий микроклимат для прорастания, накрыв плёнкой, и обеспечивают оптимальную вентиляцию. Ростки появляются примерно через месяц. На протяжении всего сезона их нужно поливать, а землю пропалывать и рыхлить. Молодые растения на зиму накрывают хвойным лапником. Уже на будущий год пихты можно перемещать на постоянное место.
Закалённые в холоде семечки высаживают в землю в апреле на глубину до 2 см, используя питательный субстрат для хвойников. Рассаде создают подходящий микроклимат для прорастания, накрыв плёнкой, и обеспечивают оптимальную вентиляцию. Ростки появляются примерно через месяц. На протяжении всего сезона их нужно поливать, а землю пропалывать и рыхлить. Молодые растения на зиму накрывают хвойным лапником. Уже на будущий год пихты можно перемещать на постоянное место.
Можно разводить пихту при помощи черенков. Для этого весной у взрослого растения отламывают однолетние веточки с верхушечной почкой и «пяточкой». Предотвратить заболевания растения можно, опустив черенки в раствор марганцовки. Чтобы обезопасить родительское дерево, срез замазывают садовым варом и заматывают полиэтиленовой плёнкой.
Чтобы укоренить веточки, необходимо:
- поместить побеги в смесь из песка, перегноя и садовой земли;
- накрыть стеклом и поставить в тепло, каждый день проводя проветривание;
- на зимний период убрать в прохладное, сухое место, например, в подвал или погреб;
- весной контейнеры выносят на улицу, а в осеннее время их уже можно посадить в открытый грунт.

Наиболее простой метод – размножение отводками, когда здоровые нижние побеги растения пригибают к почве и фиксируют скобой в подготовленную борозду, заглублённую на 5 см. Процедуру осуществляют весной и в течение всего сезона заботятся о растении, проводя полив, рыхление, удаление сорных трав и мульчирование перегноем.
В преддверии зимы отводки утепляют и закрывают, а от материнской пихты отделяют постепенно, в несколько заходов. Пересаживают новые растения спустя 1-2 года.
Болезни и вредители
Хорошо известна устойчивость хвойной культуры к разным видам вредителей и болезням, но при неграмотном уходе растение все же может заболеть. Причиной этого является избыток или застой воды. На веточках пихты появляются тёмно-коричневые пятна, а хвоя желтеет и осыпается. Это симптомы грибкового поражения – так называемой ржавчины. Для борьбы с грибком применяют опрыскивание надземной части бордоской смесью, а заражённые побеги срезают, дезинфицируя срезы садовым варом.
Причиной этого является избыток или застой воды. На веточках пихты появляются тёмно-коричневые пятна, а хвоя желтеет и осыпается. Это симптомы грибкового поражения – так называемой ржавчины. Для борьбы с грибком применяют опрыскивание надземной части бордоской смесью, а заражённые побеги срезают, дезинфицируя срезы садовым варом.
Для профилактики используют любые средства, содержащие медь, в частности, медный купорос. Обработку нужно проводить весной в конце марта и в апреле. Иногда проблему вызывает сильное загущение кроны, поэтому её следует прореживать.
Из насекомых наиболее опасным вредителем для пихты является хермес – это близкий родственник филлоксеров и тли, также паразитирующих на вечнозелёных породах деревьев. Из-за их поражения листва подвергается увяданию, пожелтению и засыханию. Предотвратить нападение насекомых можно, опрыскивая посадки раствором «Антио» (на 10 л воды нужно 20 г вещества).
Такие мероприятия актуальны после схода снега. Они помогают не допустить появления других паразитов – еловой побеговой моли, листовёртки и её личинок.
Использование в ландшафтном дизайне
Миниатюрные разновидности растения используются для озеленения скверов, парков, частных садов. Одинаково эффектно небольшие компактные деревца выглядят по отдельности и в группах. Рядом с ними рекомендуется высаживать хвойные породы – сосны, лиственницы и ели. Красиво смотрятся растения рядом с клёнами, низкорослыми декоративными и ягодными кустарниками, стелящимися почвопокровными сортами цветов.
Корейскую пихту можно располагать на газонах около беседок, искусственных прудов, композиций, элементами которых являются цветы, декоративные камни, злаковые растения. Культура подходит для оформления альпийских горок, рокариев, создания живого бордюра, причём в комбинации с другими деревьями – можжевельником, туей, кипарисом, маленькими ёлочками. Рядом с корейской пихтой и под её кроной можно высаживать тенелюбивые многолетники.
Культура подходит для оформления альпийских горок, рокариев, создания живого бордюра, причём в комбинации с другими деревьями – можжевельником, туей, кипарисом, маленькими ёлочками. Рядом с корейской пихтой и под её кроной можно высаживать тенелюбивые многолетники.
Благодаря своим размерам, декоративной хвое, зимостойкости и неприхотливости в уходе корейская пихта завоевала любовь и признание многих садоводов. Этим и объясняется спрос на любые её сорта и неизменный интерес селекционеров, продолжающих выводить новые гибридные виды с необыкновенными характеристиками.
Посмотреть на красоту пихты корейской можно далее.
Пихта корейская: декоративные формы и сорта
Пихта корейская впервые была найдена сто лет назад на юге Корейского полуострова.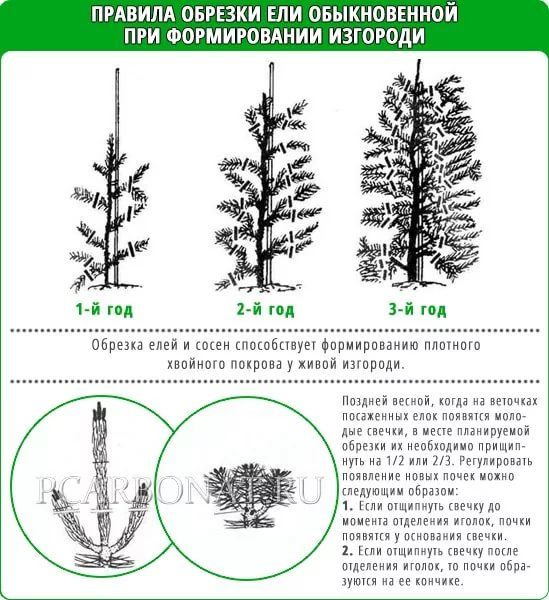
В гористых местностях, забираясь на высоту до 1800 м над уровнем моря, она образует как смешанные, так и чистые заросли. В природе – это дерево 15–18 м высотой. Растёт очень медленно, годовой прирост не превышает 4–5 см. Крона красивая, правильная, ширококоническая. Хвоя – блестящая, тёмно-зелёная, снизу – голубовато-белая. Плодоносить начинает в очень раннем возрасте. Шишки цилиндрические, до 6–7 см длиной, вначале сине-фиолетовые, затем, созревая, становятся коричневато-пурпурные; созревают в конце лета – начале осени.
Пихта корейская отличается высокой зимостойкостью. Зона 4. Имеет множество культиваров.
Adelba (Адельба) – карликовая форма, едва достигающая 1,0–1,5 в высоту; диаметр у кроны равен или чуть больше высоты.
Растёт очень медленно, годовой прирост не превышает 2–3 см. Скелетные побеги короткие, толстые, умеренно ветвящиеся, разрастаются от центра растения выражено неравномерно, отходят под острым углом и направлены в стороны и вверх. В результате формируется неправильная, асимметричная, широкая, уплощенно-овальная или подушковидная крона. Хвоя – короткая, плоская, с закруглённым кончиком, она густо покрывает побеги и окрашена в тёмно-зелёные цвет, снизу хвоинки – голубовато-белые. Цвет молодых приростов резко отличаются от тональности старой хвои, они окрашены в яркий травянисто-зелёный тон. Растение можно посадить как на открытое, солнечное место, так и в лёгкую тень; уровень освещённости на декоративные характеристики не влияет. Почвы желательны питательные, умеренно увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение, благодаря своим небольшим размерам может успешно использоваться в садах любой площади. Оно украсит альпийские горки, каменистые, вересковые и японские сады, подпорные стенки, прогулочные дорожки, прибрежные зоны водоёмов, опушки леса. Долгое время может жить в контейнерах.
В результате формируется неправильная, асимметричная, широкая, уплощенно-овальная или подушковидная крона. Хвоя – короткая, плоская, с закруглённым кончиком, она густо покрывает побеги и окрашена в тёмно-зелёные цвет, снизу хвоинки – голубовато-белые. Цвет молодых приростов резко отличаются от тональности старой хвои, они окрашены в яркий травянисто-зелёный тон. Растение можно посадить как на открытое, солнечное место, так и в лёгкую тень; уровень освещённости на декоративные характеристики не влияет. Почвы желательны питательные, умеренно увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение, благодаря своим небольшим размерам может успешно использоваться в садах любой площади. Оно украсит альпийские горки, каменистые, вересковые и японские сады, подпорные стенки, прогулочные дорожки, прибрежные зоны водоёмов, опушки леса. Долгое время может жить в контейнерах.
Aurea (Ауреа) – карликовая, кустообразная форма.
Высота взрослого растения 1,0–1,3 м; диаметр у кроны около полутора метров. Растёт медленно, годовой прирост не превышает 5–6 см. Скелетные побеги разной длины, отходят от центра растения под прямым углом и лежат в горизонтальной плоскости, разрастаются в стороны выражено неравномерно, сильно ветвятся. Крона формируется неправильная, асимметричная, в молодом возрасте уплощенно-округлая, с возрастом – характерная, гнездовидная. Хвоя – короткая, очень широкая, с закруглённым кончиком, насыщенно-зелёного цвета, нижняя сторона хвоинок – светло-голубая или голубовато-белая. Молодые приросты в лёгкой тени – лимонно-жёлтые, на освещённых местах становятся ярко-травянисто-зелеными. Солнечными ожогами хвоя не страдает. Растение желательно высаживать в притененное место, это даёт возможность добиться более яркой окраски хвои. Почвы необходимы питательные, увлажнённые, но без застоя воды, дренированные. Зимостойкость хорошая. Зона 4. Это карликовое растение поможет внести оживление в оформление альпийской горки, каменистого и верескового сада, подпорной стенки прогулочной дорожки, причём его можно высаживать в места любой экспозиции.
 Может расти в контейнерах.
Может расти в контейнерах.
Blue Cones (Блю Коунс) – относительно высокорослое дерево, достигает 8–9 м в высоту, диаметр у кроны около 3,5–4,0 м. Растёт медленно, годовой прирост составляет 5–7 см, не более. Ствол ровный, прямой. Скелетные ветви толстые, прямые, от ствола отходят практически под прямым углом, разрастаются равномерно, сильно ветвятся. Концы веток и боковые побеги слегка приподняты и направлены вверх. Крона очень красивая, правильная, симметричная, плотная, пирамидальная или широкопирамидальная. Хвоя – толстая, широкая, длинная, синевато-зелёная с серебристым оттенком, снизу – голубовато-белая. Дерево рано вступает в плодоношение, и его долго украшают многочисленные пурпурно-фиолетовые шишки. Созревая, они становятся темно-коричневато-фиолетовыми. Растение можно посадить как на открытое, солнечное место, так и в тенистое; уровень освещённости на декоративности дерева не сказывается. Почвы необходимы питательные, увлажнённые, но без застоя воды, дренированные. Зимостойкость хорошая. Зона 4. Такое нарядное дерево станет украшением парадной зоны сада, его можно посадить в виде солитера на газоне, опушке лесной зоны. Зимой – это отличная новогодняя ёлка.
Зимостойкость хорошая. Зона 4. Такое нарядное дерево станет украшением парадной зоны сада, его можно посадить в виде солитера на газоне, опушке лесной зоны. Зимой – это отличная новогодняя ёлка.
Blue Standard (Блю Стандард) – высокорослое дерево, своим габитусом мало отличающееся от исходного вида. Высота взрослого растения достигает 12–15 м, а диаметр у кроны около 4,0 м. Растёт относительно быстро, прибавляя ежегодно до 8–10 см. Скелетные ветви длинные, толстые, сильно ветвящиеся, разрастаются в стороны равномерно, отходят от прямого, ровного ствола под почти прямым углом и направлены в бок и слегка вверх. Крона формируется плотная, правильная, симметричная, ширококоническая или пирамидальная. Хвоя – блестящая, длинная, до 4,0–4,5 см длиной, толстая, с закруглёнными кончиками, серповидная; окрашена в тёмно-зелёный цвет, снизу хвоинки – голубовато-белые. Примечательны шишки этого сорта, сначала они сине-пурпурные, затем становятся интенсивно-сине-фиолетовыми. При создании этого сорта селекция и шла в направлении отбора растения с более тёмной, насыщенной окраской шишке, нежели у других культиваров. Растение может расти как на солнечном, так и тенистом месте. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость высокая. Зона 4. Дерево с такими внушительными габаритами в основном рассчитано на сады больших площадей, но и в маленьком саду ему можно отвести парадное место или роль солитера на газоне; растение может участвовать в формировании лесного участка, свободно растущей живой изгороди, из него получается великолепная новогодняя ёлка.
Растение может расти как на солнечном, так и тенистом месте. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость высокая. Зона 4. Дерево с такими внушительными габаритами в основном рассчитано на сады больших площадей, но и в маленьком саду ему можно отвести парадное место или роль солитера на газоне; растение может участвовать в формировании лесного участка, свободно растущей живой изгороди, из него получается великолепная новогодняя ёлка.
Blauer Pfiff (Блауэр Рфиф) – карликовая, скорее кустообразная форма, едва достигающая метра в высоту, диаметр у кроны около полутора метров.
Растёт медленно, годовой прирост не превышает 4–5 см. Скелетные ветви разной длины, большая их часть – короткие, разрастаются в стороны выражено неравномерно, от центра отходят под углом и направлены в стороны и вверх. Крона формируется неправильная, асимметричная, округло-подушковидная. Хвоя – сравнительно короткая, очень толстая, прямая, густо расположенная на побегах, окрашена в насыщенный сине-зелёный цвет. Молодые приросты – светло-травянисто-зеленые, контрастно выделяются на фоне старой хвои. Снизу иголки голубовато-белесые. Плодоносить начинает очень рано; шишки несколько меньшего размера, чем у исходного вида, многочисленные, густо-сине-фиолетовые. Растение может расти как на освещённых, солнечных местах, так и в затенении, на декоративных качествах уровень освещённости не сказывается. Почвы желательные питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение для оформления альпийской горки, каменистого или верескового сада, подпорной стенки, опушки леса, прогулочной дорожки; может быть выразительным солитером на газоне.
Молодые приросты – светло-травянисто-зеленые, контрастно выделяются на фоне старой хвои. Снизу иголки голубовато-белесые. Плодоносить начинает очень рано; шишки несколько меньшего размера, чем у исходного вида, многочисленные, густо-сине-фиолетовые. Растение может расти как на освещённых, солнечных местах, так и в затенении, на декоративных качествах уровень освещённости не сказывается. Почвы желательные питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение для оформления альпийской горки, каменистого или верескового сада, подпорной стенки, опушки леса, прогулочной дорожки; может быть выразительным солитером на газоне.
Brevifolia (Бревифолиа) – карликовая, кустообразная форма, достигающая полутора метров в высоту; диаметр у кроны равен или чуть больше высоты.
Растёт медленно, годовой прирост не превышает 4–5 см. Скелетные ветви короткие, толстые, умеренно ветвящиеся, разрастаются в стороны равномерно. В молодом возрасте побеги отходят от центра под углом и направлены в стороны и вверх; крона округлая. С возрастом центральный побег несколько вытягивается, и отчётливо намечается верхушка; крона становится приземистой, широкопирамидальной. В любом возрасте она плотная, густоветвистая. Хвоя – короткая, толстая, изогнутая, насыщенно-зелёная, снизу – пепельно-белая. Шишки – мелкие, многочисленные, фиолетовые. Растение успешно может расти на освещённых, солнечных и в тенистых местах. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение для оформления альпийской горки, каменистого и верескового сада, подпорной стенки, склона, прибрежной зоны водоёма, прогулочной дорожки, лесного уголка сада.
С возрастом центральный побег несколько вытягивается, и отчётливо намечается верхушка; крона становится приземистой, широкопирамидальной. В любом возрасте она плотная, густоветвистая. Хвоя – короткая, толстая, изогнутая, насыщенно-зелёная, снизу – пепельно-белая. Шишки – мелкие, многочисленные, фиолетовые. Растение успешно может расти на освещённых, солнечных и в тенистых местах. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение для оформления альпийской горки, каменистого и верескового сада, подпорной стенки, склона, прибрежной зоны водоёма, прогулочной дорожки, лесного уголка сада.
Brilliant (Брильянт) – невысокая, кустообразная форма, достигающая 1,0–1,2 м в высоту; диаметр у кроны около метра.
Растёт медленно, годовой прирост 5–6 см. Скелетные ветви сильно ветвящиеся, короткие, разной длины, от центра растения отходят хаотично, под углом и направлены в стороны и вверх, разрастаются неравномерно. Крона формируется плотная, густоветвистая, округлая, асимметричная. Хвоя – блестящая, короткая, тонкая, плоская, насыщенно-зелёная, снизу – голубовато-белая. Молодые приросты – травянисто-зелёные, контрастно выделяются на фоне старой хвои. Шишки мелкие, многочисленные, сине-фиолетовые. Растение может расти как на освещённых, солнечных местах, так и в затенении. Почвы необходимы питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Нарядное, яркое растение для украшения альпийской горки, каменистого и верескового сада, подпорной стенки, склона, опушки леса, свободно растущей живой изгороди, прогулочной дорожки как на открытом месте, так и лесной зоне, берега водоёма; прекрасный солитер на газоне. Может расти в контейнере.
Хвоя – блестящая, короткая, тонкая, плоская, насыщенно-зелёная, снизу – голубовато-белая. Молодые приросты – травянисто-зелёные, контрастно выделяются на фоне старой хвои. Шишки мелкие, многочисленные, сине-фиолетовые. Растение может расти как на освещённых, солнечных местах, так и в затенении. Почвы необходимы питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Нарядное, яркое растение для украшения альпийской горки, каменистого и верескового сада, подпорной стенки, склона, опушки леса, свободно растущей живой изгороди, прогулочной дорожки как на открытом месте, так и лесной зоне, берега водоёма; прекрасный солитер на газоне. Может расти в контейнере.
Compact Dwarf (Компакт Дворф) – карликовая кустообразная форма.
Высота взрослого растения не более 70–80 см в высоту; диаметр у кроны около полутора метров. Растёт относительно быстро, годовой прирост составляет 8–10 см, в благоприятные годы до 12 см. Скелетные ветви сильно ветвящиеся, разрастаются в стороны равномерно, большая их часть – длинные, отходящие от центра под прямым углом и в бок, лежат в горизонтальной плоскости, некоторые – на земле. Некоторые, более укорочённые побеги в верхней части растения отходят от центра под углом и направлены в стороны и вверх. Крона формируется плотная, густая, правильная, симметричная, в молодом возрасте округлая, с возрастом начинает уплощаться и становится подушковидной. Хвоя – короткая, толстая, прямая, густо сидящая на побегах; окрашены в густой зелёный цвет, молодые приросты светлее на тон. Снизу хвоинки голубовато-белые. Растение лучше сажать на открытые или слегка затенённые места, в значительной тени побеги вытягиваются, и крона становится рыхловатой. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость высокая. Зона 4. Прекрасное растение для оформления склонов, подпорных стенок, опушек леса, альпийских горок, каменистых и вересковых садов, прогулочных дорожек.
Некоторые, более укорочённые побеги в верхней части растения отходят от центра под углом и направлены в стороны и вверх. Крона формируется плотная, густая, правильная, симметричная, в молодом возрасте округлая, с возрастом начинает уплощаться и становится подушковидной. Хвоя – короткая, толстая, прямая, густо сидящая на побегах; окрашены в густой зелёный цвет, молодые приросты светлее на тон. Снизу хвоинки голубовато-белые. Растение лучше сажать на открытые или слегка затенённые места, в значительной тени побеги вытягиваются, и крона становится рыхловатой. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость высокая. Зона 4. Прекрасное растение для оформления склонов, подпорных стенок, опушек леса, альпийских горок, каменистых и вересковых садов, прогулочных дорожек.
Dark Hills (Дарк Хиллз) – карликовая форма, достигающая 1,5–2,0 м в высоту, диаметр у кроны около метра.
Растёт очень медленно, годовой прирост не превышает 2–3 см. Скелетные побеги относительно короткие, разрастаются равномерно, умеренно ветвятся, от центра отходят под острым углом и направлены слегка в стороны и вверх. Крона формируется плотная, округлая, симметричная. Хвоя – матовая, длинная, плоская, толстая, густо расположенная на побегах, с закруглённым кончиком. Она окрашена в насыщенно-зелёный цвет, снизу хвоинки голубовато-белые. Молодые приросты – ярко-травянисто-зеленые, контрастно выделяются на фоне старой хвои. Растение можно сажать как на освещённые солнцем места, так и в затенение. Уровень освещённости на декоративности растения не сказывается. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение можно посадить в каменистый, вересковый и японский сад, на альпийскую горку, подпорную стенку, склон, опушку леса, включить в состав свободно растущей живой изгороди.
Крона формируется плотная, округлая, симметричная. Хвоя – матовая, длинная, плоская, толстая, густо расположенная на побегах, с закруглённым кончиком. Она окрашена в насыщенно-зелёный цвет, снизу хвоинки голубовато-белые. Молодые приросты – ярко-травянисто-зеленые, контрастно выделяются на фоне старой хвои. Растение можно сажать как на освещённые солнцем места, так и в затенение. Уровень освещённости на декоративности растения не сказывается. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение можно посадить в каменистый, вересковый и японский сад, на альпийскую горку, подпорную стенку, склон, опушку леса, включить в состав свободно растущей живой изгороди.
Doni-Tajuso (Дони Таджусо) – карликовая кустообразная форма.
Высота растения около полуметра, диаметр у кроны чуть больше высоты. Растёт медленно, прибавляя ежегодно 2–3 см. Скелетные ветви короткие, сильно ветвящиеся, плотно прижатые друг к другу, разрастаются в стороны равномерно, от центра растения отходят под углом и направлены в стороны и вверх. Крона формируется очень плотная, густая, симметричная, уплощенно-шаровидная или округлая. Растение напоминает слегка приплюснутый шар. Хвоя – длинная, до 3,0–3,5 см длиной, относительно тонкая, серповидная, тёмно-зелёная, снизу пепельно-голубая. Молодые приросты – сочно-зелёные; ярко выделяются на фоне старой хвои. Растение может расти на участках сада любой экспозиции, уровень освещённости на декоративность не влияет. Почвы необходимы умеренно питательные и увлажнённые, дренированные. Зимостойкость высокая. Зона 4. Прекрасное растение для оформления альпийской горки, каменистого, верескового и японского сада, подпорной стенки, прибрежной зоны водоёма, прогулочных дорожек. Для усиления декоративного эффекта растения высаживают небольшими группами. Может расти в контейнере.
Крона формируется очень плотная, густая, симметричная, уплощенно-шаровидная или округлая. Растение напоминает слегка приплюснутый шар. Хвоя – длинная, до 3,0–3,5 см длиной, относительно тонкая, серповидная, тёмно-зелёная, снизу пепельно-голубая. Молодые приросты – сочно-зелёные; ярко выделяются на фоне старой хвои. Растение может расти на участках сада любой экспозиции, уровень освещённости на декоративность не влияет. Почвы необходимы умеренно питательные и увлажнённые, дренированные. Зимостойкость высокая. Зона 4. Прекрасное растение для оформления альпийской горки, каменистого, верескового и японского сада, подпорной стенки, прибрежной зоны водоёма, прогулочных дорожек. Для усиления декоративного эффекта растения высаживают небольшими группами. Может расти в контейнере.
Felderback (Фельдербак) – карликовая кустообразная форма.
Высота взрослого растения около полутора метров; диаметр у кроны равен или чуть больше высоты. Растёт медленно, годовой прирост не превышает 4–5 см. Скелетные побеги короткие, толстые, умеренно ветвящиеся, разрастаются в стороны выражено неравномерно, отходят от центра под углом и направлены в стороны и вверх. Крона – неправильная, асимметричная, плотная, округлая, с возрастом становится уплощенно-округлой. Хвоя – блестящая, тонкая, длинная, со слегка заострённым кончиком, очень густо располагается на побегах. Окрашена в насыщенный сине-зелёный цвет; нижняя сторона хвоинок – голубоватая. Молодые приросты – ярко-травянисто-зеленые, контрастно выделяются на фоне старой хвои. Шишки – некрупные, темно-пурпурно-фиолетовые. Растение лучше высаживать на открытые места, в затенении побеги нередко излишне вытягиваются, и крона становится рыхловатой. Почвы необходимы питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Яркое украшение альпийской горки, каменистого, верескового или японского сада, подпорной стенки, склона, опушки леса, растение можно включить в состав свободно растущей живой изгороди.
Скелетные побеги короткие, толстые, умеренно ветвящиеся, разрастаются в стороны выражено неравномерно, отходят от центра под углом и направлены в стороны и вверх. Крона – неправильная, асимметричная, плотная, округлая, с возрастом становится уплощенно-округлой. Хвоя – блестящая, тонкая, длинная, со слегка заострённым кончиком, очень густо располагается на побегах. Окрашена в насыщенный сине-зелёный цвет; нижняя сторона хвоинок – голубоватая. Молодые приросты – ярко-травянисто-зеленые, контрастно выделяются на фоне старой хвои. Шишки – некрупные, темно-пурпурно-фиолетовые. Растение лучше высаживать на открытые места, в затенении побеги нередко излишне вытягиваются, и крона становится рыхловатой. Почвы необходимы питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Яркое украшение альпийской горки, каменистого, верескового или японского сада, подпорной стенки, склона, опушки леса, растение можно включить в состав свободно растущей живой изгороди.
Fliegente Untertasse (Флайгенте Унтертассе) – карликовое деревце или кустообразная форма.
Достигает 1,5–2,0 м в высоту, диаметр у кроны равен высоте. Растёт сравнительно быстро, годовой прирост 7–10 см. Скелетные ветви толстые, короткие, сильно ветвящиеся, особенно на концах, разрастаются в стороны равномерно, отходят от центрального побега под острым углом и направлены в стороны и вверх. Крона формируется правильная, симметричная, округлая, с возрастом в центре образуется небольшое углубление, и крона превращается в гнездовидную. Хвоя – длинная, тонкая, со слегка заострённым кончиком, серповидно изогнутая, насыщенного зелёного цвета, нижняя сторона хвоинок серебристо-белая. Молодые приросты светлее на тон. Растение можно сажать на открытые места, в лёгкую тень и, даже в значительное затенение. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Растение можно посадить на альпийскую горку, в каменистый, вересковый или японский сад, на подпорную стенку, склон, опушку леса, включить в состав свободно растущей живой изгороди.
Green Carpet (Грин Карпит) – карликовая кустообразная форма.
Высота взрослого растения не более полуметра, диаметр у кроны от метра до полутора. Растёт относительно быстро, прибавляя ежегодно 8–10 см. Скелетные ветви толстые, длинные, разрастаются в стороны выражено неравномерно, отходят от центра растения под почти прямым углом и направлены широко в стороны. Концы побегов лежат на земле. Крона формируется неправильная, асимметричная, в молодом возрасте – округлая, с годами уплощается и становится подушковидной, широко раскинутой в стороны. Хвоя – длинная, плоская, широкая, с закруглённым кончиком, насыщенного зелёного цвета. Молодая хвоя – ярко-травянисто-зеленая, выделяется на фоне старой хвои, но не резко. Растение одинаково хорошо растёт на освещённых, солнечных местах, и в затенении. Почвы желательны питательные, рыхлые, умеренно увлажнённые. Зимостойкость хорошая. Зона 4. Место растения на альпийской горке, в каменистом, вересковом или японском саду, на подпорной стенке и склоне, опушке леса и вдоль прогулочной дорожки. Оно может исполнять роль великолепного почвенного покрытия в местах любой экспозиции.
Оно может исполнять роль великолепного почвенного покрытия в местах любой экспозиции.
Luminetta (Люминетта) – карликовое деревце или кустообразная форма.
Высота взрослого растения 1,5–2,0 м; диаметр у кроны около метра. Растёт сравнительно быстро, годовой прирост составляет7–8 см. Скелетные ветви толстые, прямые, сильно ветвящиеся, разрастающиеся в стороны равномерно; они отходят от центра растения под углом и направлены в бок и вверх. Крона формируется симметричная, правильная, плотная, уплощенно-округлая. Хвоя – блестящая, длинная, толстая, с закруглённым кончиком, густо расположенная на побегах, окрашена в насыщенный зелёный цвет; нижняя сторона хвоинок – светло-серебристо-белая. Молодые приросты – яркие, травянисто-зелёные. Шишки – некрупные сине-пурпурные. Растение можно посадить на освещённое солнцем место или в лёгкую, сквозистую тень. Почвы желательны питательные, увлажнённые, но без застоя воды. Зимостойкость хорошая. Зона 4. Растение для каменистых, вересковых и восточных садиков, больших альпийских горок, украшения подпорных стенок, склонов, опушек леса. Его можно высадить одиночно на газоне, а также включить в состав свободно растущей живой изгороди.
Его можно высадить одиночно на газоне, а также включить в состав свободно растущей живой изгороди.
Molli (Молли) – красивое, стройное дерево высотой 5–7 м; диаметр у кроны около 2 м.
Растёт сравнительно медленно, годовой прирост не превышает 6–7 см. Ствол ровный, прямой. Скелетные ветви – толстые, прямые, от ствола отходят под небольшим углом и направлены в стороны и вверх, разрастаются равномерно, сильно ветвятся, особенно на концах. Крона формируется плотная, правильная, симметричная, ширококоническая или пирамидальная с чётким выразительным силуэтом. Хвоя – средней длины, широкая, плоская, густо расположенная на побегах, окрашена в насыщенный зелёный цвет с лёгким синеватым оттенком; снизу хвоинки нежно-голубовато-белые. Молодые приросты – ярко травянисто-зелёные, контрастно выделяются на фоне старой хвои. Шишки очень крупные, многочисленные, синие с фиолетовым оттенком.
Хотя растение теневыносливо, его лучше высаживать на открытых, освещённых солнцем местах. В условиях затенения побеги излишне вытягиваются, и крона становится рыхловатой. Почвы требуются питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Это нарядное дерево украсит парадную зону сада, может быть участником смешанной группы, формирующей свободно стоящую живую изгородь, станет эффектным солитером на газоне, а зимой новогодней ёлкой.
В условиях затенения побеги излишне вытягиваются, и крона становится рыхловатой. Почвы требуются питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Это нарядное дерево украсит парадную зону сада, может быть участником смешанной группы, формирующей свободно стоящую живую изгородь, станет эффектным солитером на газоне, а зимой новогодней ёлкой.
Piccolo (Пикколо) – карликовая кустообразная форма.
Высота взрослого растения 30–35 см; диаметр у кроны около полутора метров. Растёт сравнительно быстро, годовой прирост равен 8–10 см. Скелетные ветви длинные, толстые, сильно ветвящиеся, особенно на концах, разрастаются неравномерно. У молодых растений побеги отходят под небольшим углом и направлены слега вверх и стороны. Крона формируется уплощенно-овальная. С возрастом побеги отходят от центра растения под почти прямым углом и располагаются горизонтально. Крона становится уплощенной, широко раскидистой. В любом возрасте она достаточно плотная, асимметричная. Хвоя – длинная, плоская, широкая, с закруглённым кончиком, густо расположенная на побегах. Она окрашена в насыщенный зелёным цвет; нижняя сторона – серебристо-белая. Молодые приросты – ярко-травянисто-зеленые; они контрастно выделяются на фоне старой хвои. Дерево можно посадить как на открытое, освещённое солнцем место, так и в лёгкую тень, декоративность растения при этом не страдает. Почва подходит умеренно питательная, увлажнённая, дренированная. Зимостойкость хорошая. Зона 4. Изящное почвопокровное растение можно использовать для озеленения альпийской горки, каменистого, верескового или японского садика, подпорных стенок, склонов, опушек леса, прогулочных дорожек, берегов водоёма.
Хвоя – длинная, плоская, широкая, с закруглённым кончиком, густо расположенная на побегах. Она окрашена в насыщенный зелёным цвет; нижняя сторона – серебристо-белая. Молодые приросты – ярко-травянисто-зеленые; они контрастно выделяются на фоне старой хвои. Дерево можно посадить как на открытое, освещённое солнцем место, так и в лёгкую тень, декоративность растения при этом не страдает. Почва подходит умеренно питательная, увлажнённая, дренированная. Зимостойкость хорошая. Зона 4. Изящное почвопокровное растение можно использовать для озеленения альпийской горки, каменистого, верескового или японского садика, подпорных стенок, склонов, опушек леса, прогулочных дорожек, берегов водоёма.
Silberlocke (Зильберлоке) – невысокое, скорее – карликовое, стройное дерево, достигающее высоты 1,5–2,0 м; диаметр у кроны около полутора метров.
Растёт медленно, годовой прирост составляет 6–8 см. Ствол ровный, прямой. Скелетные ветви прямые, ровные, сильно ветвящиеся, равномерно разрастающиеся в стороны; от ствола они отходят под небольшим углом и направлены в стороны и вверх. Крона – симметричная, правильная, ширококоническая или пирамидальная, с чётким красивым силуэтом. Иногда формируется две — три вершины. Хвоя очень необычная. Она – длинная, плоская, густо расположенная на побеге, закрученная внутрь, сверху она окрашена в тёмно-зелёный цвет, снизу – серебристо-белая. Благодаря такому строению хвоинок побеги кажутся серебристо-зелёными.
Крона – симметричная, правильная, ширококоническая или пирамидальная, с чётким красивым силуэтом. Иногда формируется две — три вершины. Хвоя очень необычная. Она – длинная, плоская, густо расположенная на побеге, закрученная внутрь, сверху она окрашена в тёмно-зелёный цвет, снизу – серебристо-белая. Благодаря такому строению хвоинок побеги кажутся серебристо-зелёными.
Растение лучше желательно сажать на открытые, солнечные места, при таком местоположении крона формируется плотная, компактная. Почвы необходимы питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Дерево настолько яркое, нарядное, имеющее совершенно необычный облик, что его сажают таким образом, чтобы обеспечить круговой обзор. Это может быть парадная зона сада, одиночная посадка на газоне, подпорной стенке, опушке леса или склоне.
Silberzwerg (Зильберцверг) – карликовое дерево, достигающее полутора метров в высоту, диаметр у кроны равен или чуть больше высоты.
Растёт медленно; годовой прирост равен 3–5 см. Скелетные ветви длинные, толстые, сильно ветвящиеся, от ствола отходят под острым углом и направлены в стороны и вверх, разрастаются равномерно. Крона симметричная, правильная, плотная, широкопирамидальная с чётким правильным силуэтом. Хвоя – длинная, толстая, плоская, слегка серповидная, тёмно-зелёная, иногда с лёгким синеватым оттенком. Снизу хвоинки – голубовато-белые. Молодые приросты ярко-травянисто-зеленые; контрастно выделяются на фоне старой хвои. Растению желательно подобрать открытое, солнечное местоположение, в затенённых местах побеги излишне вытягиваются, и крона становится рыхловатой. Почвы требуются питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Это карликовое деревце можно посадить одиночно на газоне, альпийской горке, каменистом, вересковом и японском саду, подпорной стенке, склоне, опушке леса. Оно может выступить в роли новогодней ёлки.
Verdener Dom (Верденер Дом) – карликовое деревце, высотой около 2,5 м; диаметр у кроны 80–90 см.
Растёт медленно, годовой прирост не превышает 3–5 см. Ствол ровный, прямой. Скелетные ветви – короткие, толстые, сильно ветвящиеся, равномерно разрастающиеся в стороны, от ствола отходят под небольшим углом и направлены в стороны и вверх. Крона формируется плотная, густая, симметричная, коническая, с чётким силуэтом. Хвоя – короткая, толстая, прямая, радиально отстоящая от побега, расположена очень густо, окрашена в насыщенный зелёный цвет. Нижняя сторона хвоинок серебристо-белая. Молодые приросты на тон светлее. Шишки – сине-фиолетовые. Дерево может расти как на открытых местах, так и в значительном затенении, уровень освещённости на декоративности растения не сказывается. Почвы желательны питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Небольшое деревце с плотным, компактным габитусом способно украсить альпийскую горку, каменистый, вересковый и японский садик, подпорную стенку, склон, опушку леса, войти в состав смешанной группы древесных растений, формирующих свободно растущую живую стенку.
Veredlung (Вередлунг) – карликовая кустообразная форма.
Высота взрослого растения около полутора метров; диаметр у кроны равен или чуть больше высоты. Растёт довольно быстро, прибавляя ежегодно до 8–10 см. Скелетные ветви толстые, сильно ветвящиеся, от центра растения отходят под небольшим углом, сначала направляются в стороны и вверх, затем, слегка дуговидно изгибаясь, ложатся горизонтально, разрастаются побеги выражено неравномерно. Крона – неправильная, асимметричная, плотная, овальная; с возрастом в центре формируется небольшое гнездовидное углубление. Хвоя – блестящая, длинная, толстая, плоская, с закруглённым кончиком, серповидно изогнутая, окрашена в насыщенный зелёный цвет. Обратная сторона хвоинок – пепельно-белая. Молодые приросты – яркие, травянисто-зелёные; они контрастно выделяются на фоне старой хвои. Растению подбирают открытое, освещённое солнцем место, в затенении побеги излишне вытягиваются, и крона становится рыхлой. Почвы необходимы питательные, увлажнённые, дренированные. Зимостойкость хорошая. Зона 4. Отличное растение для оформления альпийской горки, каменистого, верескового или японского сада, подпорной стенки, склона, прогулочной дорожки, прибрежной зоны, опушки леса.
Зимостойкость хорошая. Зона 4. Отличное растение для оформления альпийской горки, каменистого, верескового или японского сада, подпорной стенки, склона, прогулочной дорожки, прибрежной зоны, опушки леса.
Автор: Н. Кузнецова
Правила прищипывания ели, сосны, можжевельника, других хвойных растений
Мы научились управлять ростом лиственных деревьев и кустарников, формируя их с помощью стрижки. А можно ли то же самое делать с хвойными растениями? Оказывается, можно. Но инструмент используем другой – прищипывание молодых побегов. Когда и как проводить прищипку хвойных, рассказываем в статье.
Прищипка – один из способов формирования густой и пышной кроны у хвойных растений
Cледите за нами:
Зачем нужна прищипка
Скажем сразу, прищипка хвойных пород – вовсе не обязательная процедура. Растение прекрасно чувствует себя и без нее, формируя крону естественных очертаний. Но бывают ситуации, когда без внешнего вмешательства не обойтись.
В каких случаях используют прищипку?
- Когда нужно сдержать рост дерева в высоту и/или ширину.
- Если из-за неравномерного развития веток в кроне появились проплешины и это место нужно загустить.
- Если растение пострадало во время зимы (сломанные ветки, деформированная крона).
- Чтобы сохранить силуэт растения, поддержать и усилить его густоту.
- Чтобы предупредить неправильное формирование кроны у молодых, недавно высаженных растений.
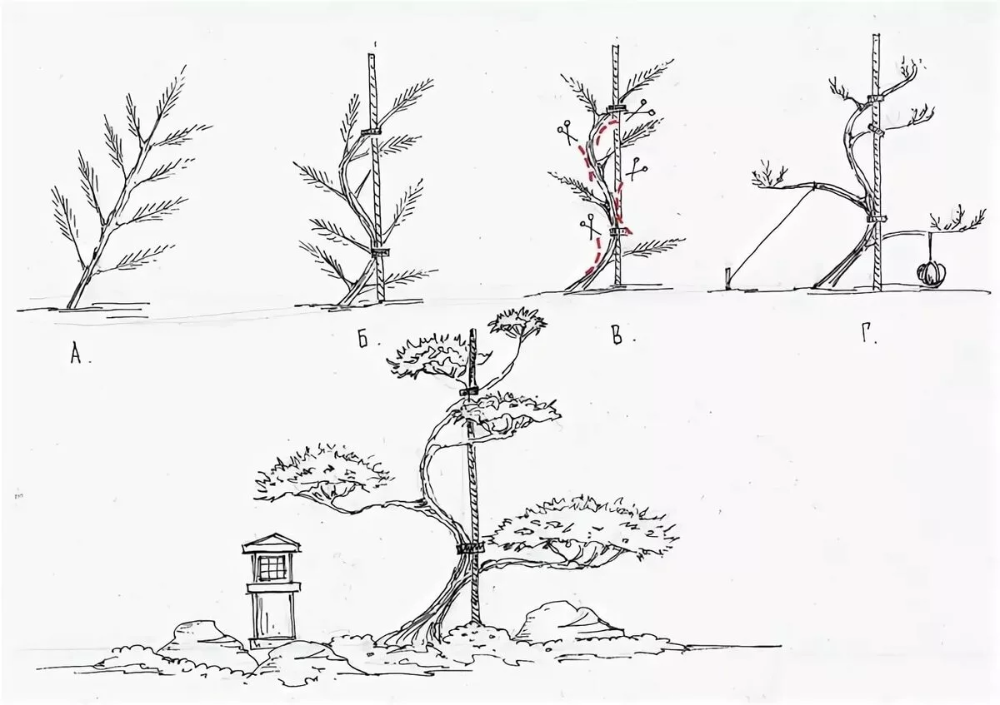
Прищипка – один из основных методов выращивания карликовых деревьев бонсай. Но искусство бонсай – это целая система, которая, помимо прищипывания молодых побегов, включает и другие приемы: обрезку взрослых ветвей, выбор почек, выщипывание хвои, а также подвязывание и растяжку веток.
Виды прищипки
- Сплошное прищипывание – равномерное укорачивание всех молодых побегов по аналогии со стрижкой лиственных растений.
- Удаление в каждой мутовке центрального проводника – вместо одной ветки продолжения вырастет несколько и хвойная лапа получится более пышная.

- Прищипка самых длинных приростов – можно управлять ростом веток и перенаправлять энергию с более сильных побегов на слабые.
Чтобы держать крону такой сосны в форме, достаточно ежегодно прищипывать молодые приросты
Общие правила прищипки хвойных растений
- Хвойные можно начинать прищипывать в любом возрасте. К примеру, ель достигла 1,5-метровой высоты и вы хотите, чтобы она осталась в этих размерах. Или вам достался участок с уже взрослыми растениями, которым нужно придать опрятную форму. Но если вы изначально хотите получить карликовую ель, прищипку нужно начинать как можно раньше.
- Оптимальное время для прищипки хвойных – когда молодые побеги полностью сформированы, но хвоя еще не раскрылась. У разных пород эти сроки немного отличаются, но в среднем приходятся на конец весны – начало лета.
- Чтобы получить аккуратную и симметричную крону, прищипывать нужно все побеги – и главные, и боковые.
- Сколько прищипывать, зависит от декоративных задач.
 Обычно рекомендуют удалять не менее 1/3 побега, а оставлять – не менее 1/4.
Обычно рекомендуют удалять не менее 1/3 побега, а оставлять – не менее 1/4. - Побеги лучше не обрезать, а выламывать, чтобы не повредить хвою на будущей макушке ветки. Иголки живут от 2 до 5 лет, и если вы их наполовину срежете, они не отрастут и все это время будут выглядеть неопрятно.
- Начав прищипку хвойных, желательно делать ее каждый сезон. Если год-два пропустите процедуру, декоративный эффект пропадет.
Прежде чем начинать прищипку хвойных, оцените объем работ и свою готовность выполнять это ежегодно
Особенности прищипывания в зависимости от породы
Хвойные включают несколько видов растений с разными типами хвои и моделями роста. Рассмотрим сроки и технологию прищипки наиболее распространенных пород.
Сосна обыкновенная и горная
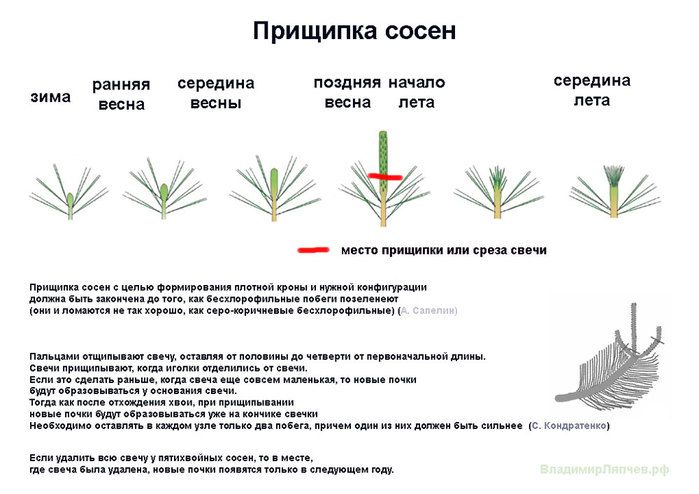
У сосен ярко выраженный верхушечный тип роста. Центральный проводник всегда вырастает из почки, находящейся на макушке побега, от него в сторону отходит еще 3–4 прироста, образуя мутовку.
Молодые побеги, внешне похожие на свечки, отрастают к концу весны. Их рекомендуют обламывать в тот момент, когда побег уже освободился от чешуек, но иголки еще остаются прижатыми.
В этот период почки на следующий год еще не заложены – они начнут формироваться чуть позже, когда раскроется хвоя. И если вовремя растение прищипнуть, оно будет воспринимать место надлома как новую макушку.
Укоротить нужно все «свечки»: самые крупные – сильнее, слабые – поменьше. Чем короче побеги вы оставите, тем гуще и пышнее получится крона.
Места прищипки ничем замазывать не нужно. На месте надлома выступит капелька смолы, которая сама обеззаразит и затянет ранку.
Оптимальное время для прищипывания сосны определяют по состоянию «свечек»
Ель обыкновенная и голубая
У елей молодые побеги растут по-другому. Основное их количество, как и у сосен, сосредоточено на макушке, но часть приростов разбросана по всей длине ветки.
Считается, что побег готов к прищипке, когда его размер достигает 1–2 см, но хвоя еще не раскрылась. По календарю это время приходится на май – начало июня.
По календарю это время приходится на май – начало июня.
Вы можете использовать два способа укорачивания побегов:
- прищипнуть каждый из них;
- обрезать часть ветки с приростом, но следить, чтобы на оставшейся части были молодые побеги, которые ее продолжат.
У ели сильнее укорачивают приросты на макушке и развитых ветках, меньше – в нижней части и внутри кроны
Кедр
Кедр прищипывают, если нужно приостановить рост основного побега и стимулировать образование боковых веточек. Для этого удаляют точку роста на макушке. Делают это весной – в апреле-мае.
Если дерево плохо приживается, медленно растет, болеет, к прищипке лучше не прибегать.
Так выглядит молодой побег кедра, рост которого можно остановить, прищипнув макушку
Туя, можжевельник
Стрижку и прищипку туй, можжевельников и других растений с чешуйчатой хвоей делают, чтобы сохранить контур и загустить крону.
Поскольку эти породы активно растут весь сезон, стричь и прищипывать их можно весной и летом. Лучше всего это делать в начале лета, когда все почки распустились и хорошо видно молодую хвою. Вторую стрижку проводят в августе, когда молодые побеги начинают одревесневать. Этим вы стимулируете ветвление на следующий год.
Лучше всего это делать в начале лета, когда все почки распустились и хорошо видно молодую хвою. Вторую стрижку проводят в августе, когда молодые побеги начинают одревесневать. Этим вы стимулируете ветвление на следующий год.
У можжевельника для сдерживания роста достаточно прищипнуть молодые приросты и перенаправить тем самым энергию растения на развитие боковых побегов.
В отличие от других хвойных, туи активно наращивают чешуйчатые листья весь сезон
Щипать или не щипать хвойные
Среди садоводов немало противников прищипки хвойных.
Главным недостатком этого приема они считают слишком большие временные затраты, которые с каждым годом увеличиваются. Вначале это может быть несколько десятков побегов, но спустя годы они превратятся в сотни, а то и тысячи.
Второй минус – обязательность процедуры. Если уж начали щипать, будьте добры делать это каждый год, иначе весь труд пойдет насмарку, а растение через несколько лет превратится в неопрятный «веник».
Если у вас нет опыта прищипки хвойных, мы рекомендуем начать с одного-двух растений. Это поможет понять технику, набраться опыта, оценить временные затраты. И самостоятельно сделать вывод – щипать или не щипать.
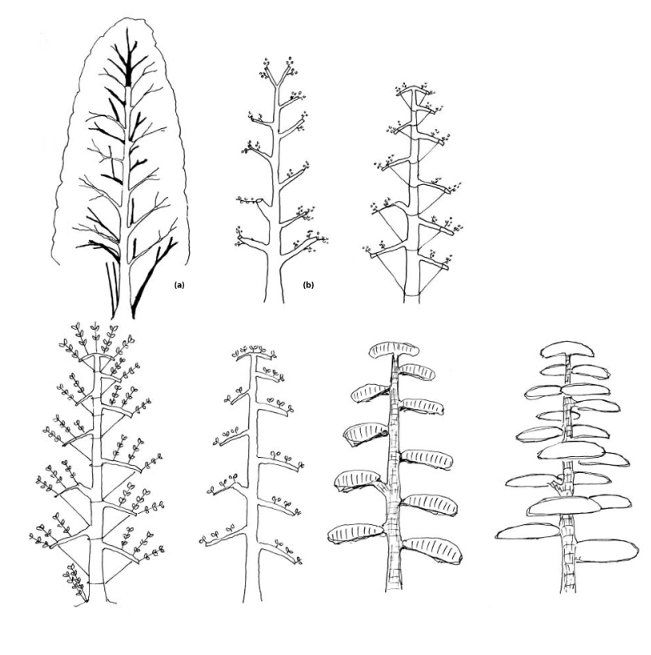
Строение кроны и типы побегов пихты
Урожай семян у древесных растений — результат сложного взаимодействия всех частей кроны и, следовательно, в значительной степени зависит от ее строения. Поэтому изучение вопросов плодоношения пихты сибирской проведено нами с учетом особенностей расположения различных побегов в кроне дерева.
Исследования строения кроны проводились на срубленных деревьях в возрасте от 17 до 140 лет. В кронах деревьев подсчитывалось количество мутовок, основных и межмутовочных ветвей, измерялись межмутовочные промежутки и длина ветвей. В каждой мутовке и межмутовочном промежутке отмечались возраст и пол составляющих ветвь побегов (годичных приростов). Пол побегов у пихты сибирской легко определить по следам генеративных органов: на женских побегах отчетливо видны стержни от рассыпавшихся шишек, длительное время остаются различимыми следы и на мужских побегах.

Сведения, полученные при детальном анализе строения кроны, позволяют судить не только о количественном соотношении разных типов побегов в кроне, но и. раскрывают особенности их качественных изменений (длительность женской сексуализации, смена пола), а также воссоздают картину неравномерности урожаев.
Пирамидальная крона пихты сибирской отличается мутовчатым ветвлением, с большим количеством межмутовочных побегов. Верхние ветви обычно растут под острым углом к оси дерева, средние — под прямым, а нижние вследствие значительной длины сильно отклоняются от ствола и свисают к земле. В густых насаждениях нижние ветви из-за недостатка освещения часто отмирают и кроны деревьев оказываются несколько поднятыми над землей.
Кроны деревьев у пихты сибирской значительно отличаются друг от друга своей густотой, обусловленной различным содержанием межмутовочных ветвей. В качестве объективного признака густокронности можно использовать среднюю нагрузку одного метра оси дерева межмутовочными ветвями. У деревьев с типичной кроной она не превышает 50 ветвей, а у густокронных достигает 89.
У деревьев с типичной кроной она не превышает 50 ветвей, а у густокронных достигает 89.
Большая разница наблюдается и в количестве межмутовочных ветвей второго порядка. Для сравнения приведем данные о нагрузке ветвей межмутовочными побегами второго порядка на участке кроны последнего десятилетия. У типичных деревьев на одной ветви образуется в среднем от 0,7 до 3,5 межмутовочных побегов, в то время как у густокронных нагрузка ветвей — от 3,0 до 4,7 побега.
Некоторое совмещение этих показателей кроется в заведомо разном количестве ветвей первого порядка у типичных и густокронных деревьев, в результате нагрузка на ветви несколько выравнивается. В качестве наглядного примера количественной разницы межмутовочных побегов второго порядка можно привести абсолютное количество их у разных деревьев. Так, у типичных деревьев на участке кроны последнего десятилетия оно не превышает 250, а у густокронных достигает 654 побегов.
Такое строение кроны у деревьев пихты сибирской нельзя считать случайным, поскольку крона дерева представляет собой определенную систему, в которой каждый элемент играет особую роль.
Пихта сибирская, как и все представители семейства сосновых, — однодомное растение. Характерная черта цветения — раздельнополые стробилы, располагающиеся на побегах в разных частях кроны.
В связи с этим у пихты наблюдается явная разнокачественность побегов, проявляющаяся типом их сексуализации. Так же как у сосны обыкновенной и кедра сибирского, у пихты наблюдается три типа побегов: ростовые, сложные ростовые + женские и ростовые + мужские.
Наиболее простые по своему строению — ростовые побеги: удлиненные, почти гладкие, с редкими волосками и спирально сидящей мягкой хвоей. На большинстве побегов хвоя скручена у основания и расположена настильно. В наиболее затененной части кроны она располагается в два ряда (гребенчато). Заканчивается ростовой побег терминальной почкой, обеспечивающей рост в следующем году.
Сложные побеги совмещают в себе ростовой стержень с хвоей и видоизмененными побегами следующего порядка в виде микро — и мегастробилов, несущих уже генеративные функции. На этом основании некоторые авторы выделяют микростробилы и женские шишки в самостоятельные типы побегов.
На этом основании некоторые авторы выделяют микростробилы и женские шишки в самостоятельные типы побегов.
Сложные ростовые + женские (кратко — женские) побеги отличаются от ростовых жесткой хвоей, отходящей в сторону почти под прямым углом, и наличием одной или нескольких вертикально расположенных женских шишек. В первый год образования такого побега на нем закладываются генеративные женские почки, из которых весной следующего года появляются шишки, созревающие осенью. Таким образом, женский побег выполняет генеративную функцию только в течение двух лет, в дальнейшем он превращается в ростовой и шишки появляются на приросте следующего года.
На нижней стороне сложных ростовых + мужских (кратко — мужских) побегов располагаются микроспорофилловые колоски от основания и в зависимости от обилия закладки до половины побега или до его вершины. Мужские побеги существуют в качестве генеративных также только два неполных года, уступая эти функции приросту следующего года.
Иногда в кроне дерева можно обнаружить обоеполые побеги, характерные для семейства сосновых, однако на таких побегах получают полное развитие генеративные зачатки обычно только одного пола.
Побеги каждого типа занимают характерное место в кроне дерева. Женские побеги появляются на хорошо развитых ветвях в верхней части кроны, образуя женский генеративный ярус, который занимает от 4 до 10 верхних мутовок и имеет протяженность 1,4—2,2 м, иногда до 3,5 м.
Мужской ярус занимает 10—20 мутовок в средней части кроны. 1—2 мутовки между женским и мужским генеративными ярусами представляют смешанный ярус. Здесь на наиболее сильных побегах образуются шишки, а более слабые несут микроспорофилловые колоски. Как правило, основные ветви смешанного генеративного яруса еще образуют женские побеги, а большинство межмутовочных ветвей, даже расположенных гораздо выше по кроне, бывают уже полностью покрыты мужскими побегами.
Нижняя часть кроны представлена только ростовыми побегами. Ростовой ярус обычно имеет протяженность около 5—10 м.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Как правильно стричь елку — Статьи
Сроки обрезки хвойных растений зависят от поставленных целей и их особенностей роста. Плановую обрезку проводят как весной, так и осенью. Во время весенней обрезки удаляют сухие и часть старых побегов. Для того, чтобы ограничить рост растений, проводят прищипку молодых побегов. Осенняя процедура призвана не только сдержать прирост кроны, убрать остатки переболевших ветвей, на которых могут оставаться источники заражения, но и лучше подготовиться к зимнему периоду.
Плановую обрезку проводят как весной, так и осенью. Во время весенней обрезки удаляют сухие и часть старых побегов. Для того, чтобы ограничить рост растений, проводят прищипку молодых побегов. Осенняя процедура призвана не только сдержать прирост кроны, убрать остатки переболевших ветвей, на которых могут оставаться источники заражения, но и лучше подготовиться к зимнему периоду.
Вынужденная обрезка проводится по необходимости – в любое время года, как только проявятся признаки заболевания или результаты нашествия насекомых-вредителей.
Формирующую обрезку весной можно совместить с плановой, если во время последней удалено лишь небольшое количество ветвей. Чтобы стресс для растения свести к минимуму, проводить формирование необходимо до распускания почек. В течение лета каждые 2 – 3 недели можно проводить коррекцию кроны, удаляя выросшие более других, торчащие веточки. Осенью, вместе с плановой, проводится последняя формовка кроны.
Сроки топиарной обрезки определяют исходя из физиологических особенностей растения, но до выхода растения из зимней спячки. Исключением являются сосны, обрезку которых проводят в мае-июне, после появления молодых приростов, до образования новой хвои. После придания растению нужной формы в течение всего сезона поводят необходимую стрижку, обрезку или прищипывание побегов.
Исключением являются сосны, обрезку которых проводят в мае-июне, после появления молодых приростов, до образования новой хвои. После придания растению нужной формы в течение всего сезона поводят необходимую стрижку, обрезку или прищипывание побегов.
При работе с хвойными растениями необходимо знать их особенности. Например, стрижку ели лучше проводить тогда, когда происходит закладка почек — во второй половине июня. В зависимости от сорта им можно придавать очень разные формы. Большинство елей имеют форму кроны в виде правильного конуса, особенно хороши ели колючие и их сорта Blue Diamond, Edith, Fat Albert, Hoopsii, Iseli Fastigiate, Koster, Omega и многие другие. Такую форму легко подчеркнуть, сдержать в определенных размерах или сформировать из неё колонну. Существуют ели в виде шаров и подушек разной величины – из них легко сформировать шары. Ель обыкновенная, несмотря на быстрый рост, хорошо формируется в геометрические фигуры (шары, кубы, конусы, живые изгороди). Ель обыкновенная Нифидормис несет крону в виде подушки, и из нее со временем можно сформировать зонтик на вытянутой ножке. Густая крона ели сизой Коника может превратиться почти в идеальный медленнорастущий конус.
Ель обыкновенная Нифидормис несет крону в виде подушки, и из нее со временем можно сформировать зонтик на вытянутой ножке. Густая крона ели сизой Коника может превратиться почти в идеальный медленнорастущий конус.
Формирование кроны сосны включает и обрезку – прореживание кроны, и укорачивание – прищипку свежих свечек, и придание ветвям определённой формы с помощью растяжек. Таким образом сосне можно придать форму шара и позже только поддерживать этот образ, прищипывая молодой прирост. Живописные сосны очень часто используются для создания ниваки. Прореживание кроны и жесткий каркас помогают создать неповторимые формы.
Молодой прирост сосен и елей боится возвратных заморозков, поэтому работу с ним лучше начинать не ранее второй половины мая. Прищипка верхушечного прироста останавливает рост растения вверх, а бокового – возможность создания густой и пышной кроны, так как будет закладываться не одна, а несколько боковых почек. Результат работы будет виден через 2 – 3 года.
Формировать крону лиственницы лучше ранней весной, до того, как почки проснутся. Плановая обрезка осенью поможет подготовится к зиме. Растение очень пластичное, и с помощью постоянных прищипок можно не только поддерживать силуэт растения, но и сдерживать её рост.
Можжевельники плохо переносят осеннюю стрижку, и поэтому формированием кроны лучше заниматься с весны до середины августа. Древесина можжевельника – плотная, каждый срез – это тяжелая травма, и поэтому следует не тянуть, и срезать не уже одревесневшие ветви, а молодой, зеленый прирост.
Стричь пихту и тую хорошо в периоды неактивного движения смолы – ранней весной или в июне, учитывая естественный рост кроны при формировании силуэта.
Вопрос: Как посадить пихту? — Растения
Содержание статьи:
Как вырастить пихту из семян, Часть 1.
 Сбор семян.
Сбор семян.Видео взято с канала: Очумелый садовод
Хвойные растения в саду Пихта
Видео взято с канала: Марина Марти
ПИХТА корейская Сильберлок/ Посадка и Уход/ Формирование кроны
Показать описаниеВыращивание пихты корейской имеет особенности, которые связаны с климатом, составом почвы. На примере пихты корейской Сильберлок расскажу об основном уходе за пихтой, посадка пихты, почва для пихты, кислотность грунта при посадке пихты корейской, полив пихты, мульчирование приствольного круга..
Покажу как формирую пихту Сильберлок. С помощью прищипки свечейприростов пихты можно формировать крону, сделать ее гуще, направить в определенную сторону рост, контролировать разрастание. Уже взрослые пихты высотой более 1.5 м легче и быстрее стричь кусторезом или мотоножницами, хотя есть недостатки такой обрезки травмируется хвоя пихты. Чем обработать пихту после обрезки, как формировать верхушку пихты, как выглядит пихта Сильберлок в мае покажу в видео. .
.
Красавица пихта корейская сорт Сильберлок удивит интересным окрасом хвои, декоративностью и неприхотливостью, а вырастить Пихту в саду, зная основные моменты агротехники Под силу каждому!
На канале в плейлисте Любимые хвойные собраны видео о видах и сортах хвойных растений https://www.youtube.com/playlist?list=PLNKbMv103Ncx3m2us9jwzw688g8SJus3p.
Плейлист Хвойные растения https://www.youtube.com/playlist?list=PLNKbMv103NczS2DkLECO6H53pKKiV7tmI.
информация о хвойных деревьях и кустарниках в нашем саду, уход в разное время года, стрижка, обрезка хвойных, подкормки, обработки, вредители и болезни хвойных..
Подписывайтесь на канал Под силу каждому!
https://www.youtube.com/c/Подсилукаждому.
#пихта #хвойные #подсилукаждому #пихтасильберлок #стрижкапихты #хвойныйсад.
#пихтакорейская #stihl
Видео взято с канала: Под силу каждому!
пересадка пихты
Видео взято с канала: Экзоты Крыма
Пересадка пихты
Видео взято с канала: Обо всем по-немногу
Посадка пихты как правильно посадить пихту
Видео взято с канала: Саженцы от питомника “Добрый Сад”
Пихта / посадка пихты / уход за пихтой
Видео взято с канала: О РАСТЕНИЯХ
Как посадить пихту на даче. Перед посадкой пихты нужно понимать, что это хвойное растение – долгожитель. Для него прожить 300 лет и более – вообще не проблема. Как правильно посадить пихту. Для посадки пихты предпочтительно выбирать пасмурные, теплые дни, а лучше всего сажать пихту под дождь.
Как посадить пихту: пошаговая инструкция с фото. Пересаживать саженцы из контейнеров на постоянное место можно в любое время. Как посадить пихту.
За две недели до того, как сажать пихту, выкопайте яму размером примерно 60х60х60, хотя габариты ямы зависят от объема корневой системы саженца. Пихта в горшке: как вырастить пихту в домашних условиях. Если саженец куплен в зимнюю пору, то перед тем как посадить пихту в постоянный горшо. 2 Как вырастить пихту на даче: посадка, выращивание, уход и виды пихт 2.1 Как посадить пихту на даче 2.2 Какая почва лучше. Как посадить пихту черенком в открытый грунт.
На следующий год укоренившиеся растения переносят в открытый грунт. Для пихты выбирают солнечное место или полутень. Пихта теневынослива, поэтому для ее посадки подойдет как тенистое, так и солнечное место. Однако следует помнить, дерево приобретает. Как я сажала пихту Мне очень повезло: есть у меня подружка Ленка с немалым опытом в садовом деле. – Пихтовые саженцы покупай старше 4 лет, а высаживать их нужно в мае или в сентябре.
Пихта теневынослива, поэтому для ее посадки подойдет как тенистое, так и солнечное место. Однако следует помнить, дерево приобретает. Как я сажала пихту Мне очень повезло: есть у меня подружка Ленка с немалым опытом в садовом деле. – Пихтовые саженцы покупай старше 4 лет, а высаживать их нужно в мае или в сентябре.
2 Как посадить пихту. 2.1 Подготовка посадочной Как размножить пихту. Существует 3 основных способа – посев семян, черенкование и использование отводков.
Пихта может украсить любую аллею или живую изгородь. Инструкция по посадке пихты, как правильно посадить и ухаживать за пихтой. Как посадить пихту из семян. Посадку семян дерева лучше проводить в конце марта, в рыхлую супесчаную почву поверх первого слоя опилок, затем засыпать и.
Процедура установки зубных коронок
Обычно требуется не менее двух посещений стоматолога для подготовки зуба к установке зубной коронки. Целью первого визита к стоматологу является осмотр и подготовка зуба, на который будет установлена коронка, и создание слепка зуба для изготовления коронки. Во время этого визита также обычно устанавливается временная коронка для защиты зуба.
Во время этого визита также обычно устанавливается временная коронка для защиты зуба.
Во время второго визита, который происходит после того, как зуботехническая лаборатория изготовила коронку, можно снять временную коронку и зафиксировать постоянную изготовленную на заказ коронку.
Иллюстрация процедуры установки зубной коронки. Изображение предоставлено: BlueRingMedia / Shutterstock
Первый визит: осмотр, изменение формы зуба, оттиск и временная коронка
Во время первого визита к стоматологу обычно делается рентген зуба и челюстной кости для выявления признаков кариеса и других факторов, которые могут увеличить риск инфицирования или повреждения пульпы зуба. Некоторым пациентам может потребоваться лечение корневых каналов перед установкой коронки на зуб.
Зубу необходимо придать форму, чтобы было место для установки коронки; степень изменения формы будет зависеть от типа используемой коронки. Металлические коронки имеют тенденцию быть тоньше и, следовательно, требуют удаления меньшего количества тканей зуба, чем фарфоровые коронки. Верхняя и боковые стороны зуба обтачиваются, чтобы придать зубу форму, необходимую для коронки. Любой кариес также следует лечить на этом этапе.
Верхняя и боковые стороны зуба обтачиваются, чтобы придать зубу форму, необходимую для коронки. Любой кариес также следует лечить на этом этапе.
Что такое идеальная зубная коронка Play
У некоторых пациентов с тяжелым кариесом или повреждением зуба может быть недостаточно оставшейся ткани зуба для установки зуба.В этом случае может потребоваться пломбировочный материал для создания структуры зуба, чтобы впоследствии можно было установить коронку.
После того, как зубу будет придана адекватная форма, следующим шагом будет создание оттиска сформированного зуба, чтобы можно было сделать коронку, максимально точно подходящую к зубу. Также часто необходимо создать оттиск для зуба на противоположной челюсти, который соединится с коронкой, когда пациент прикусывает. Это делается для того, чтобы коронка хорошо прилегала к другой стороне рта, в дополнение к области, где она будет сидеть.
Слепки обычно изготавливаются с использованием пасты или замазки, удерживающей форму зуба. Он распределяется по копируемой области, а затем отрывается, сохраняя на своей поверхности форму соответствующих зубов.
Он распределяется по копируемой области, а затем отрывается, сохраняя на своей поверхности форму соответствующих зубов.
На этом этапе важно записать сведения о зубе, которые повлияют на изготовление коронки, например, естественную посадку и цвет. В идеале цвет коронки должен совпадать с цветом зуба и окружающих его зубов, чтобы его не заметили, когда он находится в полости рта.
Наконец, на зуб обычно устанавливается временная коронка, чтобы защитить его до тех пор, пока коронка не будет изготовлена и готова к фиксации на месте. Его удалят при следующем посещении стоматолога, чтобы освободить место для постоянной коронки.
Второй визит: Установка постоянной коронки
Тем временем слепки зубов отправляются зубному технику или в лабораторию для формирования коронки правильной формы. Этот процесс обычно занимает до трех недель, поэтому для правильной установки постоянной коронки требуется более одного визита к стоматологу.
После того, как зубной техник или лаборатория изготовили коронку, наступает время установки коронки на зуб. Если установлена временная коронка, ее сначала следует снять.
Если установлена временная коронка, ее сначала следует снять.
Перед цементированием важно убедиться, что коронка изготовлена правильно, чтобы посадка и цвет коронки соответствовали цвету естественных и окружающих зубов.
Обычно вводят местный анестетик, чтобы обезболить область во время процедуры установки коронки.Стоматологический цемент используется для фиксации коронки на зубе. Может потребоваться внести небольшие коррективы в форму коронки, особенно если есть проблемы с прикусом пациента, когда коронка соприкасается с противоположным зубом.
Процедура установки коронок в тот же день
В некоторых случаях коронки могут изготавливаться в стоматологической практике, чтобы их можно было изготовить и установить в тот же день, без необходимости отправки оттисков в зуботехническую лабораторию.
Вместо того, чтобы делать оттиск зуба, для фотографирования зубов используется сканирующее устройство, называемое палочкой.Эти изображения загружаются в компьютерную программу, которая создает 3D-модель зуба. Затем эта цифровая модель может быть использована для изготовления керамической коронки в тот же день. Этот метод известен как компьютерное проектирование и изготовление зубных коронок (CAD/CAM), и его можно выполнить примерно за 15 минут, чтобы можно было быстро установить коронку на место.
Затем эта цифровая модель может быть использована для изготовления керамической коронки в тот же день. Этот метод известен как компьютерное проектирование и изготовление зубных коронок (CAD/CAM), и его можно выполнить примерно за 15 минут, чтобы можно было быстро установить коронку на место.
Каталожные номера
Дополнительное чтение
Влияние живой кроны на вертикальные модели плотности и роста древесины пихты Дугласа.
Влияние живой кроны на вертикальные модели плотности и роста древесины пихты Дугласа. | Поиск по дереву Перейти к основному содержанию .gov означает, что это официально.
Веб-сайты федерального правительства часто заканчиваются на .gov или .mil. Прежде чем делиться конфиденциальной информацией, убедитесь, что вы находитесь на сайте федерального правительства.
Сайт защищен.
https:// гарантирует, что вы подключаетесь к официальному веб-сайту и что любая предоставленная вами информация шифруется и передается безопасно.
Автор(ы):
Барбара Л. Гартнер
Эрик М.Север
Г.Р. Джонсон
Райан Синглтон
Тип публикации:
Научный журнал (JRNL)
Первичная(ые) станция(и):
Тихоокеанская северо-западная исследовательская станция
Источник:
Канадский журнал лесных исследований. 32: 439-447
32: 439-447
Описание
С экономической точки зрения было бы полезно знать, каковы биологические триггеры образования зрелой древесины (в настоящее время высокой ценности) и (или) что поддерживает производство ювенильной древесины (в настоящее время малоценной), для разработки лесохозяйственных режимов, контролирующих относительную продуктивность из двух пород дерева. Лесоводы обычно предполагают, что ствол хвойных пород дает молодую древесину в кроне и зрелую древесину под ней.Мы проверили это предположение, сравнив площадь и ширину годичных колец и компоненты плотности древесины трех внешних годичных колец в дисках, взятых из разных вертикальных положений 34-летней пихты Дугласа ( Pseudotsuga menziesii (Mirb.) Franco). 18 деревьев были отобраны с одного участка и имели широкий диапазон высоты живой кроны. Наибольшая дисперсия (63–93%) в характеристиках древесины (площадь годичных колец: общая, ранняя древесина, поздняя древесина; ширина годичных колец: общая, ранняя древесина, поздняя древесина; доля поздней древесины: по площади, ширине; и плотность годичных колец: общая, ранняя древесина, поздняя древесина) была обусловлена различиями внутри дерева (связанными с возрастом диска). Ступенчатый регрессионный анализ дал нам уравнения для оценки характеристик древесины, после чего мы проанализировали остатки с помощью линейной модели, которая учитывала, находится ли диск внутри или ниже кроны (определяется как самый нижний узел на стволе с менее чем тремя живыми ветвями). После поправки на положение дерева и диска только 2–10% остаточной вариации были связаны с тем, находился ли диск в живой кроне или вне ее. Не было статистически значимых различий при p = 0,05 между данным диском (по номеру узла) в сравнении с вне коронки ни по одному из изученных факторов.При этом характеристики плотности древесины не были статистически значимыми при р = 0,30. Это исследование показывает, что положение кроны не влияло на переход от ювенильной к зрелой древесине, судя по плотности древесины. Таким образом, мы не нашли доказательств в поддержку концепции о том, что расстояние между деревьями и обрезка живых ветвей оказывают значительное влияние на камбиальный возраст перехода от ювенильной к зрелой древесине у деревьев дугласовой пихты этого возраста.
Ступенчатый регрессионный анализ дал нам уравнения для оценки характеристик древесины, после чего мы проанализировали остатки с помощью линейной модели, которая учитывала, находится ли диск внутри или ниже кроны (определяется как самый нижний узел на стволе с менее чем тремя живыми ветвями). После поправки на положение дерева и диска только 2–10% остаточной вариации были связаны с тем, находился ли диск в живой кроне или вне ее. Не было статистически значимых различий при p = 0,05 между данным диском (по номеру узла) в сравнении с вне коронки ни по одному из изученных факторов.При этом характеристики плотности древесины не были статистически значимыми при р = 0,30. Это исследование показывает, что положение кроны не влияло на переход от ювенильной к зрелой древесине, судя по плотности древесины. Таким образом, мы не нашли доказательств в поддержку концепции о том, что расстояние между деревьями и обрезка живых ветвей оказывают значительное влияние на камбиальный возраст перехода от ювенильной к зрелой древесине у деревьев дугласовой пихты этого возраста.
Цитата
Гартнер, Барбара Л.; Норт, Эрик М .; Джонсон, Г. Р.; Синглтон, Райан. 2002. Влияние живой кроны на вертикальные модели плотности и роста древесины пихты Дугласа. Канадский журнал лесных исследований. 32: 439-447
Примечания к публикации
- Мы рекомендуем вам также распечатать эту страницу и прикрепить ее к распечатке статьи, чтобы сохранить полную информацию о цитировании.
- Эта статья была написана и подготовлена У.S. Государственные служащие в служебное время и, следовательно, находятся в открытом доступе.
https://www.fs.usda.gov/treesearch/pubs/4926
%PDF-1.3
%
1 0 объект
>
эндообъект
7 0 объект
>
эндообъект
2 0 объект
>
ручей
2014-02-06T18:10:54+05:30Adobe Acrobat 9.02014-02-07T15:46:18+05:302014-02-07T15:46:18+05:30Adobe Acrobat 9. 0 Paper Capture Plug-inapplication/pdfuuid:00e88739 -e4cc-4bf6-afb3-b3b5e52719cbuuid:84de5550-271c-4714-8901-005639271e7c конечный поток
эндообъект
3 0 объект
>
эндообъект
4 0 объект
>
эндообъект
5 0 объект
>
эндообъект
6 0 объект
>
эндообъект
8 0 объект
>
/Шрифт >
/ProcSet [/PDF /Text /ImageB /ImageC /ImageI]
/XОбъект >
>>
/Повернуть 0
/Большой палец 33 0 R
/TrimBox [0 0 453.
0 Paper Capture Plug-inapplication/pdfuuid:00e88739 -e4cc-4bf6-afb3-b3b5e52719cbuuid:84de5550-271c-4714-8901-005639271e7c конечный поток
эндообъект
3 0 объект
>
эндообъект
4 0 объект
>
эндообъект
5 0 объект
>
эндообъект
6 0 объект
>
эндообъект
8 0 объект
>
/Шрифт >
/ProcSet [/PDF /Text /ImageB /ImageC /ImageI]
/XОбъект >
>>
/Повернуть 0
/Большой палец 33 0 R
/TrimBox [0 0 453. 6 680,28]
/Тип /Страница
>>
эндообъект
9 0 объект
>
эндообъект
10 0 объект
>
эндообъект
11 0 объект
>
эндообъект
12 0 объект
>
эндообъект
13 0 объект
>
ручей
х+
6 680,28]
/Тип /Страница
>>
эндообъект
9 0 объект
>
эндообъект
10 0 объект
>
эндообъект
11 0 объект
>
эндообъект
12 0 объект
>
эндообъект
13 0 объект
>
ручей
х+
Границы | Реакция растений на Agrobacterium tumefaciens и развитие корончатого галла
Введение
Agrobacterium tumefaciens вызывает болезнь корончатого галла у широкого круга видов-хозяев путем переноса и интеграции части своей собственной ДНК, Т-ДНК, в геном растения (Chilton et al., 1977). Этот уникальный способ действия также сделал бактерию важным инструментом в селекции растений.После прикрепления Agrobacterium к растительным клеткам и экспрессии множественных генов вирулентности (vir) несколько эффекторных белков вместе с Т-ДНК транспортируются в растительную клетку с помощью системы секреции IV типа (Thompson et al., 1988). ; Ward et al., 1988, 2002; Kuldau et al., 1990; Shirasu et al., 1990; Beijersbergen et al., 1994). Растительные факторы способствуют интеграции Т-ДНК в геном растения (Gelvin, 2000; Mysore et al. , 2000; Tzfira et al., 2004; Magori and Citovsky, 2012). После интеграции экспрессия кодируемых Т-ДНК онкогенов iaaH, iaaM и ipt индуцирует биосинтез ауксина и цитокинина (Morris, 1986; Binns and Costantino, 1998).Повышение уровня этих фитогормонов приводит к усилению пролиферации и образованию корончатых галлов. Несмотря на перенос бактериальных белков в растительную клетку, большинство штаммов Agrobacterium не вызывают реакции гиперчувствительности (ГР), связанной с быстрой и локальной гибелью клеток (Staskawicz et al., 1995). Такая реакция часто возникает, когда растения заражены бактериальными патогенами, и служит для ограничения роста и распространения патогенов на другие части растения.Соответственно, системная резистентность широкого спектра действия по всему растению (системная приобретенная резистентность, SAR) не индуцируется. В течение первых нескольких часов совместного культивирования пути защиты от патогенов активируются более или менее сильно в зависимости от системы растения и генотипа Agrobacterium , используемого для заражения (Ditt et al.
, 2000; Tzfira et al., 2004; Magori and Citovsky, 2012). После интеграции экспрессия кодируемых Т-ДНК онкогенов iaaH, iaaM и ipt индуцирует биосинтез ауксина и цитокинина (Morris, 1986; Binns and Costantino, 1998).Повышение уровня этих фитогормонов приводит к усилению пролиферации и образованию корончатых галлов. Несмотря на перенос бактериальных белков в растительную клетку, большинство штаммов Agrobacterium не вызывают реакции гиперчувствительности (ГР), связанной с быстрой и локальной гибелью клеток (Staskawicz et al., 1995). Такая реакция часто возникает, когда растения заражены бактериальными патогенами, и служит для ограничения роста и распространения патогенов на другие части растения.Соответственно, системная резистентность широкого спектра действия по всему растению (системная приобретенная резистентность, SAR) не индуцируется. В течение первых нескольких часов совместного культивирования пути защиты от патогенов активируются более или менее сильно в зависимости от системы растения и генотипа Agrobacterium , используемого для заражения (Ditt et al. , 2001, 2006; Veena et al., 2003; Ли и др., 2009). Защитные реакции становятся сильнее во время развития корончатого галла. Кроме того, резко меняется физиологическое поведение трансформированных клеток.В отличие от статей, в которых основное внимание уделяется молекулярному механизму, используемому бактерией для трансформации растительной клетки, здесь мы рассматриваем последние данные о реакции растения-хозяина и корончатого галла на инфекцию Agrobacterium . Особое внимание уделено роли регуляции экспрессии генов, фитогормонов и метаболизма.
, 2001, 2006; Veena et al., 2003; Ли и др., 2009). Защитные реакции становятся сильнее во время развития корончатого галла. Кроме того, резко меняется физиологическое поведение трансформированных клеток.В отличие от статей, в которых основное внимание уделяется молекулярному механизму, используемому бактерией для трансформации растительной клетки, здесь мы рассматриваем последние данные о реакции растения-хозяина и корончатого галла на инфекцию Agrobacterium . Особое внимание уделено роли регуляции экспрессии генов, фитогормонов и метаболизма.
Реакция хозяина на
Agrobacterium tumefaciens Перед переносом Т-ДНКЗащита от патогенов
Распознавание микробных патогенов играет центральную роль в индукции активных защитных реакций у растений.Консервативный флагеллиновый пептид flg22 распознается рецепторной киназой FLS2 и индуцирует экспрессию многочисленных генов, связанных с защитой, вызывая устойчивость к патогенным бактериям (Gómez-Gómez et al. , 1999, 2001; Zipfel et al., 2004; Chinchilla et al. ., 2006). Однако род Agrobacterium не может индуцировать этот тип быстрого и общего защитного ответа из-за исключительной дивергенции N-концевого консервативного домена флагеллина (Felix et al., 1999). При сравнении ранних изменений экспрессии генов после заражения вирулентным штаммом С58 Agrobacterium Agrobacterium с применением бактериального пептида elf26 (через 1 и 3 ч соответственно) становится очевидным ослабление ответов хозяина при обработке Agrobacterium .Пептид elf26, высококонсервативный мотив одного из наиболее распространенных белков микробов, распознаваемый рецепторной киназой EFR, является фрагментом фактора элонгации Tu (EF-Tu). EF-Tu запускает реакции врожденного иммунитета, связанные с устойчивостью к болезням у Arabidopsis (Kunze et al., 2004). В то время как обработка чистым elf26 индуцирует изменения экспрессии 948 генов Arabidopsis (Zipfel et al., 2006), только 35 генов индуцируются после заражения вирулентным штаммом Agrobacterium C58, что позволяет предположить, что бактерия каким-то образом нейтрализует ответ на elf26.
, 1999, 2001; Zipfel et al., 2004; Chinchilla et al. ., 2006). Однако род Agrobacterium не может индуцировать этот тип быстрого и общего защитного ответа из-за исключительной дивергенции N-концевого консервативного домена флагеллина (Felix et al., 1999). При сравнении ранних изменений экспрессии генов после заражения вирулентным штаммом С58 Agrobacterium Agrobacterium с применением бактериального пептида elf26 (через 1 и 3 ч соответственно) становится очевидным ослабление ответов хозяина при обработке Agrobacterium .Пептид elf26, высококонсервативный мотив одного из наиболее распространенных белков микробов, распознаваемый рецепторной киназой EFR, является фрагментом фактора элонгации Tu (EF-Tu). EF-Tu запускает реакции врожденного иммунитета, связанные с устойчивостью к болезням у Arabidopsis (Kunze et al., 2004). В то время как обработка чистым elf26 индуцирует изменения экспрессии 948 генов Arabidopsis (Zipfel et al., 2006), только 35 генов индуцируются после заражения вирулентным штаммом Agrobacterium C58, что позволяет предположить, что бактерия каким-то образом нейтрализует ответ на elf26. растением-хозяином (Lee et al., 2009). Следует отметить, что используемые в исследованиях экотип и возраст Arabidopsis (проросток против взрослого стебля) также могут объяснять некоторые различия в защитной реакции.
растением-хозяином (Lee et al., 2009). Следует отметить, что используемые в исследованиях экотип и возраст Arabidopsis (проросток против взрослого стебля) также могут объяснять некоторые различия в защитной реакции.
В отношении активации транскрипции генов, участвующих в ранних защитных реакциях растений, несколько исследований пришли к разным выводам. Клеточные культуры Ageratum conyzoides показали дифференциальную экспрессию защитных генов уже через 24 часа после инфицирования неонкогенным гипервирулентным штаммом Agrobacterium (Ditt et al., 2001). В суспензионных культурах табака, инфицированных различными штаммами Agrobacterium , транскрипция генов защиты увеличивалась в течение 3–6 ч, но начинала снижаться с началом переноса Т-ДНК (Veena et al., 2003). Исследование с использованием культивируемых в суспензии клеток Arabidopsis не показало изменений в уровне транскриптов в течение 4-24 часов, но активацию генов защиты через 48 часов после заражения (Ditt et al.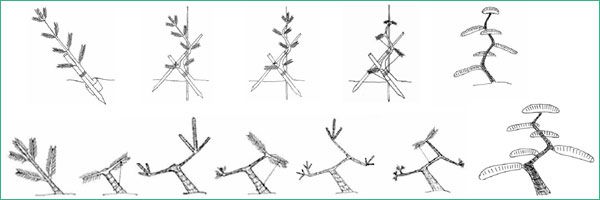 , 2006). Когда агробактерии инокулируют в основание поврежденных стеблей Arabidopsis , через 3 часа после заражения активируется очень мало защитных генов по сравнению с неинфицированными поврежденными стеблями (Lee et al., 2009). В отличие от клеточных культур, последняя экспериментальная установка не требует ни предварительной обработки фитогормонами, ни индукции гена вирулентности до заражения. Предварительная обработка фитогормонами систем клеточных культур в более ранних исследованиях может изменить защитные реакции клеток-хозяев. Таким образом, расхождения между этими исследованиями, вероятно, являются результатом использования различных систем инокуляции растений. Тем не менее, агробактерии могут злоупотреблять защитными реакциями хозяина для доставки Т-ДНК. Активируемая митогеном протеинкиназа MPK3 фосфорилирует белок VIP1 Arabidopsis , индуцируя релокализацию VIP1 из цитоплазмы в ядро.Ядерная локализация VIP1 увеличивает эффективность переноса и трансформации Т-ДНК (Djamei et al.
, 2006). Когда агробактерии инокулируют в основание поврежденных стеблей Arabidopsis , через 3 часа после заражения активируется очень мало защитных генов по сравнению с неинфицированными поврежденными стеблями (Lee et al., 2009). В отличие от клеточных культур, последняя экспериментальная установка не требует ни предварительной обработки фитогормонами, ни индукции гена вирулентности до заражения. Предварительная обработка фитогормонами систем клеточных культур в более ранних исследованиях может изменить защитные реакции клеток-хозяев. Таким образом, расхождения между этими исследованиями, вероятно, являются результатом использования различных систем инокуляции растений. Тем не менее, агробактерии могут злоупотреблять защитными реакциями хозяина для доставки Т-ДНК. Активируемая митогеном протеинкиназа MPK3 фосфорилирует белок VIP1 Arabidopsis , индуцируя релокализацию VIP1 из цитоплазмы в ядро.Ядерная локализация VIP1 увеличивает эффективность переноса и трансформации Т-ДНК (Djamei et al. , 2007).
, 2007).
Фитогормоны
Агробактерии сами продуцируют ауксин и цитокинин, чтобы модулировать реакцию растений (рис. 1А). Эти фитогормоны были определены как в клетках, так и в среде культивирования (Morris, 1986). Было высказано предположение, что биосинтез фитогормонов катализируется ферментами онкогенов, кодируемых Т-ДНК, поскольку транскрипты и белки этих генов обнаружены в клетках агробактерий (Schröder et al., 1983; Янссенс и др., 1984). Выраженные количества ауксина были определены в вирулентном штамме Agrobacterium C58, а также в более низких концентрациях в штаммах без плазмиды и с истощением Т-ДНК (Liu and Kado, 1979; Kutáeek and Rovenská, 1991). Последние данные подтвердили последние результаты (Lee et al., 2009). Тот факт, что штамм без Ti-плазмиды все еще может продуцировать ауксин, предполагает локализацию генов также вне Ti-плазмиды. Однако это предположение не подтверждается данными секвенирования штамма C58 (Wood et al., 2001). Известно, что гены, участвующие в биосинтезе ауксина, кодируются только Т-ДНК Ti-плазмиды. Недавно эти авторы определили наличие транскриптов iaaH и iaaM с помощью ПЦР в клетках Agrobacterium штамма С58 и подтвердили ранее полученные данные. Остается доказать, ответственны ли эти гены за продукцию ауксина или ауксин синтезируется по другому механизму в клетках Agrobacterium . Механизм биосинтеза цитокинина агробактериями изучен гораздо лучше.В нопалине, использующем штаммов Agrobacterium , цитокинин продуцируется в больших количествах Ti-плазмидой, кодирующей транс--зеатинсинтезирующий (tzs) фермент, ген которого расположен в регулоне vir (Akiyoshi et al., 1985, 1987; Hwang и др., 2010). Существенным меньшим источником продукции цитокинина является изопентенилированная транспортная РНК (тРНК), катализируемая хромосомно-кодируемым ферментом тРНК: изопентенилтрансферазой (MiaA), присутствующим во всех штаммах Agrobacterium (Gray et al., 1996).
Недавно эти авторы определили наличие транскриптов iaaH и iaaM с помощью ПЦР в клетках Agrobacterium штамма С58 и подтвердили ранее полученные данные. Остается доказать, ответственны ли эти гены за продукцию ауксина или ауксин синтезируется по другому механизму в клетках Agrobacterium . Механизм биосинтеза цитокинина агробактериями изучен гораздо лучше.В нопалине, использующем штаммов Agrobacterium , цитокинин продуцируется в больших количествах Ti-плазмидой, кодирующей транс--зеатинсинтезирующий (tzs) фермент, ген которого расположен в регулоне vir (Akiyoshi et al., 1985, 1987; Hwang и др., 2010). Существенным меньшим источником продукции цитокинина является изопентенилированная транспортная РНК (тРНК), катализируемая хромосомно-кодируемым ферментом тРНК: изопентенилтрансферазой (MiaA), присутствующим во всех штаммах Agrobacterium (Gray et al., 1996).
РИСУНОК 1. Реакция модельного растения Arabidopsis thaliana на Agrobacterium tumefaciens и развитие корончатого галла . (A) Вирулентные (pTi) клетки агробактерий сами продуцируют и высвобождают цитокинин и ауксин, которые повышают восприимчивость хозяина и ингибируют выработку перекиси водорода (H 2 O 2 ) и реакцию гиперчувствительности (HR) при инициации инфекции. (B) После интеграции бактериальной Т-ДНК в геном растения цитокинин и ауксин синтезируются ферментами, кодируемыми Т-ДНК, и накапливаются внутри опухоли. (C) Это вызывает массивные изменения в паттерне экспрессии генов, что приводит к метаболической и морфологической адаптации, необходимой для роста и дифференцировки опухоли. (D) Потеря воды сведена к минимуму за счет механизма защиты от стресса засухи, который вызывает повышение уровня гормона стресса АБК, и АБК-зависимой суберизации клеток для предотвращения потери воды. Испарение воды (H 2 O) с разрушенной поверхности корончатого галла приводит к поступлению воды и минералов в корончатые галлы. (E) Поскольку фотосинтез подавляется, уровни кислорода низкие, опухоль продуцирует соединения C и N гетеротрофно и получает энергию в основном анаэробно путем спиртового брожения.
(A) Вирулентные (pTi) клетки агробактерий сами продуцируют и высвобождают цитокинин и ауксин, которые повышают восприимчивость хозяина и ингибируют выработку перекиси водорода (H 2 O 2 ) и реакцию гиперчувствительности (HR) при инициации инфекции. (B) После интеграции бактериальной Т-ДНК в геном растения цитокинин и ауксин синтезируются ферментами, кодируемыми Т-ДНК, и накапливаются внутри опухоли. (C) Это вызывает массивные изменения в паттерне экспрессии генов, что приводит к метаболической и морфологической адаптации, необходимой для роста и дифференцировки опухоли. (D) Потеря воды сведена к минимуму за счет механизма защиты от стресса засухи, который вызывает повышение уровня гормона стресса АБК, и АБК-зависимой суберизации клеток для предотвращения потери воды. Испарение воды (H 2 O) с разрушенной поверхности корончатого галла приводит к поступлению воды и минералов в корончатые галлы. (E) Поскольку фотосинтез подавляется, уровни кислорода низкие, опухоль продуцирует соединения C и N гетеротрофно и получает энергию в основном анаэробно путем спиртового брожения. (F) Следовательно, развивающаяся опухоль становится метаболическим стоком для растения-хозяина, в котором накапливаются метаболиты, продуцируемые исходными листьями, и минералы, поглощаемые корнями. (G) Ауксин и цитокинин также вызывают повышение уровня этилена (ЭТ), который вместе с салициловой кислотой (СК) ингибирует вирулентность агробактерий. (H) АБК также индуцирует метилирование ДНК генома растения, тем самым регулируя экспрессию генов, чувствительных к засухе. В целом геном корончатой галлы становится гиперметилированным (Me) после инфекции Agrobacterium и, возможно, способствует сильным изменениям в экспрессии генов во время роста опухоли. Онкогены Т-ДНК остаются незатронутыми метилированием генома растения.
(F) Следовательно, развивающаяся опухоль становится метаболическим стоком для растения-хозяина, в котором накапливаются метаболиты, продуцируемые исходными листьями, и минералы, поглощаемые корнями. (G) Ауксин и цитокинин также вызывают повышение уровня этилена (ЭТ), который вместе с салициловой кислотой (СК) ингибирует вирулентность агробактерий. (H) АБК также индуцирует метилирование ДНК генома растения, тем самым регулируя экспрессию генов, чувствительных к засухе. В целом геном корончатой галлы становится гиперметилированным (Me) после инфекции Agrobacterium и, возможно, способствует сильным изменениям в экспрессии генов во время роста опухоли. Онкогены Т-ДНК остаются незатронутыми метилированием генома растения.
Более ранние исследования показали, что предварительная обработка эксплантатов либо одним ауксином, либо ауксином и цитокинином одновременно увеличивает эффективность переноса Т-ДНК и стабильную трансформацию (Krens et al., 1996; Chateau et al. , 2000), а также рост корончатого галла (Gafni et al., 1995). В этом отношении фитогормоны, продуцируемые Agrobacterium , играют роль в очень ранние моменты заражения (рис. 1А), до того, как ферменты, кодируемые Т-ДНК, катализируют синтез цитокинина и ауксина в трансформированной клетке-хозяине. Что касается механизма, вызывающего повышение восприимчивости, было высказано предположение, что фитогормоны индуцируют деление растительных клеток и что фаза клеточного цикла влияет на прикрепление агробактерий и стабильную трансформацию.Вполне вероятно, что опосредованная фитогормонами модификация физиологического состояния клетки повышает способность к трансформации и интеграции Т-ДНК. Более поздние исследования касались вопроса о молекулярном механизме и сигнальных путях, с помощью которых эти фитогормоны влияют на восприимчивость клеток-хозяев. Данные транскриптомного микрочипа через 3 часа после инокуляции Agrobacterium штамма C58 в стеблей Arabidopsis показали, что гены, которые, как известно, участвуют в зависимой от фитогормонов передаче сигналов, не индуцируются в клетках-хозяевах в этот очень ранний момент времени заражения до переноса Т-ДНК (Lee et al.
, 2000), а также рост корончатого галла (Gafni et al., 1995). В этом отношении фитогормоны, продуцируемые Agrobacterium , играют роль в очень ранние моменты заражения (рис. 1А), до того, как ферменты, кодируемые Т-ДНК, катализируют синтез цитокинина и ауксина в трансформированной клетке-хозяине. Что касается механизма, вызывающего повышение восприимчивости, было высказано предположение, что фитогормоны индуцируют деление растительных клеток и что фаза клеточного цикла влияет на прикрепление агробактерий и стабильную трансформацию.Вполне вероятно, что опосредованная фитогормонами модификация физиологического состояния клетки повышает способность к трансформации и интеграции Т-ДНК. Более поздние исследования касались вопроса о молекулярном механизме и сигнальных путях, с помощью которых эти фитогормоны влияют на восприимчивость клеток-хозяев. Данные транскриптомного микрочипа через 3 часа после инокуляции Agrobacterium штамма C58 в стеблей Arabidopsis показали, что гены, которые, как известно, участвуют в зависимой от фитогормонов передаче сигналов, не индуцируются в клетках-хозяевах в этот очень ранний момент времени заражения до переноса Т-ДНК (Lee et al. , 2009). Было показано, что индол-3-уксусная кислота (ИУК) влияет на вирулентность агробактерий, подавляя индукцию гена vir и рост агробактерий (Liu and Nester, 2006). Однако этот эффект наблюдался при относительно высоких концентрациях ауксина (25–250 мкМ). В клетках Agrobacterium общее (свободное и конъюгированное) содержание ИУК составляет 0,3 ± 0,1 мкМ, а в стеблях Arabidopsis через 3 ч после инокуляции штаммом С58 — 2,1 ± 1 мкМ, тогда как в корончатых галлах Arabidopsis содержание составляет ок. .в 10 раз выше (17,3 ± 8,8 мкМ) за счет экспрессии генов iaaH и iaaM, кодируемых Т-ДНК, и их ферментативной активности (собственные данные и Thomashow et al., 1986). Применение 1 мкМ ИУК, концентрация, обнаруженная в поврежденных и неинфицированных стеблях Arabidopsis (0,8 ± 0,2 мкМ), стимулировала рост клеток Agrobacterium , тогда как стимуляция роста исчезала при 10 мкМ и более высоких концентрациях ИУК (личное сообщение, J. Ludwig -Мюллер, Технический университет Дрездена, Германия).
, 2009). Было показано, что индол-3-уксусная кислота (ИУК) влияет на вирулентность агробактерий, подавляя индукцию гена vir и рост агробактерий (Liu and Nester, 2006). Однако этот эффект наблюдался при относительно высоких концентрациях ауксина (25–250 мкМ). В клетках Agrobacterium общее (свободное и конъюгированное) содержание ИУК составляет 0,3 ± 0,1 мкМ, а в стеблях Arabidopsis через 3 ч после инокуляции штаммом С58 — 2,1 ± 1 мкМ, тогда как в корончатых галлах Arabidopsis содержание составляет ок. .в 10 раз выше (17,3 ± 8,8 мкМ) за счет экспрессии генов iaaH и iaaM, кодируемых Т-ДНК, и их ферментативной активности (собственные данные и Thomashow et al., 1986). Применение 1 мкМ ИУК, концентрация, обнаруженная в поврежденных и неинфицированных стеблях Arabidopsis (0,8 ± 0,2 мкМ), стимулировала рост клеток Agrobacterium , тогда как стимуляция роста исчезала при 10 мкМ и более высоких концентрациях ИУК (личное сообщение, J. Ludwig -Мюллер, Технический университет Дрездена, Германия). Известно, что действие ауксина сильно зависит от дозы, стимулируя рост при низких концентрациях и ингибируя при высоких концентрациях, которые незначительно варьируются в зависимости от типа растения и ткани.Можно предположить, что при инициации инфекции относительно низкие уровни ауксина в клетках агробактерий и/или в поврежденной ткани растения стимулируют рост агробактерий, тогда как более высокие концентрации, образующиеся в корончатом галле, ингибируют вирулентность, а также рост Agrobacterium . Такой антагонистический эффект ауксина будет способствовать трансформации клетки-хозяина в начале инфекционного процесса и ингибировать вирулентность и рост агробактерий, чтобы предотвратить дальнейшую трансформацию развивающихся корончатых галлов.В отличие от ауксина, роль передачи сигналов цитокинина в восприимчивости растений хорошо известна. Недавно было показано, что цитокинин, секретируемый Agrobacterium , контролирует вирулентность посредством роста бактериальных клеток и экспрессии гена vir на ранних стадиях инфекционного процесса (Hwang et al.
Известно, что действие ауксина сильно зависит от дозы, стимулируя рост при низких концентрациях и ингибируя при высоких концентрациях, которые незначительно варьируются в зависимости от типа растения и ткани.Можно предположить, что при инициации инфекции относительно низкие уровни ауксина в клетках агробактерий и/или в поврежденной ткани растения стимулируют рост агробактерий, тогда как более высокие концентрации, образующиеся в корончатом галле, ингибируют вирулентность, а также рост Agrobacterium . Такой антагонистический эффект ауксина будет способствовать трансформации клетки-хозяина в начале инфекционного процесса и ингибировать вирулентность и рост агробактерий, чтобы предотвратить дальнейшую трансформацию развивающихся корончатых галлов.В отличие от ауксина, роль передачи сигналов цитокинина в восприимчивости растений хорошо известна. Недавно было показано, что цитокинин, секретируемый Agrobacterium , контролирует вирулентность посредством роста бактериальных клеток и экспрессии гена vir на ранних стадиях инфекционного процесса (Hwang et al. , 2010). Некоторые, но не все виды растений показали цитокинин-зависимое увеличение эффективности трансформации (Hwang et al., 2013). Цитокинин, полученный из Agrobacterium , не только действует на физиологию бактерий, но также влияет на экспрессию генов-хозяев посредством классического цитокинин-зависимого сигнального пути, включая рецепторы цитокинина и фосфотрансферный каскад (Sardesai et al., 2013). Активация этого сигнального каскада посредством цитокинина, полученного из агробактерий, приводит к ингибированию экспрессии гена транскрипционного фактора семейства Arabidopsis MYB, MTF1 (Sardesai et al., 2013). MTF1 оказался негативным регулятором восприимчивости к трансформации, блокируя экспрессию интегриноподобного белка At14a, рецептора растительной мембраны. At14A служит опорными точками для прикрепления бактерий к поверхности клетки-хозяина. Таким образом, на ранних стадиях инфекции агробактериальный ауксин и цитокинин манипулируют сигнальными путями фитогормонов растений, чтобы подготовить клетку-хозяина к трансформации.
, 2010). Некоторые, но не все виды растений показали цитокинин-зависимое увеличение эффективности трансформации (Hwang et al., 2013). Цитокинин, полученный из Agrobacterium , не только действует на физиологию бактерий, но также влияет на экспрессию генов-хозяев посредством классического цитокинин-зависимого сигнального пути, включая рецепторы цитокинина и фосфотрансферный каскад (Sardesai et al., 2013). Активация этого сигнального каскада посредством цитокинина, полученного из агробактерий, приводит к ингибированию экспрессии гена транскрипционного фактора семейства Arabidopsis MYB, MTF1 (Sardesai et al., 2013). MTF1 оказался негативным регулятором восприимчивости к трансформации, блокируя экспрессию интегриноподобного белка At14a, рецептора растительной мембраны. At14A служит опорными точками для прикрепления бактерий к поверхности клетки-хозяина. Таким образом, на ранних стадиях инфекции агробактериальный ауксин и цитокинин манипулируют сигнальными путями фитогормонов растений, чтобы подготовить клетку-хозяина к трансформации.
Помимо ауксина и цитокинина, передача сигналов защиты растений включает сеть взаимосвязанных путей, в которых салициловая кислота (SA) и жасмоновая кислота (JA) вместе с этиленом (ET) функционируют как важные сигнальные молекулы (Kunkel and Brooks, 2002). Экзогенное применение молекулы защиты растений SA к клеткам Agrobacterium ингибировало экспрессию генов vir, включая tzs, рост бактерий, прикрепление бактерий к клеткам растений и вирулентность (Yuan et al., 2007; Anand et al., 2008). Однако при инициации инфекции (через 3 ч после инфицирования) ни уровни SA, ни JA, ни гены этих сигнальных путей не повышены в тканях Agrobacterium , инфицированных Arabidopsis (Lee et al., 2009). В этот момент повышается только уровень 1-аминоциклопропан-1-карбоновой кислоты (АЦК), предшественника ЭТ, в присутствии как вирулентных, так и обезоруженных штаммов Agrobacterium , но не экспрессия маркерных генов ЭТ. -зависимый сигнальный путь защиты.Инокуляция эксплантов дыни ( Cucumis melo ) Agrobacterium также увеличивает продукцию ЭТ (Ezura et al. , 2000). Известно, что ЭТ запускает продукцию ауксинов растениями из-за повышенной экспрессии растительных генов, участвующих в биосинтезе ауксинов (Степанова и др., 2005). Ауксин повышает восприимчивость хозяина, в то время как продукция ЭТ растениями оказывает негативное влияние на вирулентность агробактерий. Применение АСС снижает опосредованный Agrobacterium перенос генов в экспланты дыни, в то время как добавление аминоэтоксивинилглицина, ингибитора АСС-синтазы, увеличивает его (Ezura et al., 2000). Снижение эффективности трансформации является результатом подавления экспрессии гена vir, но не роста Agrobacterium (Nonaka et al., 2008). Стимулирующий эффект низких концентраций ауксина на рост агробактерий и ингибирующий эффект ET на вирулентность показывает, что как Agrobacterium , так и растение-хозяин контролируют трансформацию клетки-хозяина. В совокупности на ранних стадиях инфекционного процесса цитокинин и ауксин, продуцируемые клетками Agrobacterium , оказывают стимулирующее действие на эффективность трансформации, чему частично противодействует ингибирующее действие ЭТ и СК растительного происхождения на вирулентность агробактерий.
, 2000). Известно, что ЭТ запускает продукцию ауксинов растениями из-за повышенной экспрессии растительных генов, участвующих в биосинтезе ауксинов (Степанова и др., 2005). Ауксин повышает восприимчивость хозяина, в то время как продукция ЭТ растениями оказывает негативное влияние на вирулентность агробактерий. Применение АСС снижает опосредованный Agrobacterium перенос генов в экспланты дыни, в то время как добавление аминоэтоксивинилглицина, ингибитора АСС-синтазы, увеличивает его (Ezura et al., 2000). Снижение эффективности трансформации является результатом подавления экспрессии гена vir, но не роста Agrobacterium (Nonaka et al., 2008). Стимулирующий эффект низких концентраций ауксина на рост агробактерий и ингибирующий эффект ET на вирулентность показывает, что как Agrobacterium , так и растение-хозяин контролируют трансформацию клетки-хозяина. В совокупности на ранних стадиях инфекционного процесса цитокинин и ауксин, продуцируемые клетками Agrobacterium , оказывают стимулирующее действие на эффективность трансформации, чему частично противодействует ингибирующее действие ЭТ и СК растительного происхождения на вирулентность агробактерий. Таким образом, правильный баланс фитогормонов определяет успешность заражения.
Таким образом, правильный баланс фитогормонов определяет успешность заражения.
Сверхчувствительный ответ
Изучение ранних событий патогенеза показало, что вирулентная Agrobacterium не индуцирует HR у Arabidopsis (рис. 1А; Lee et al., 2009). Более того, Agrobacterium способен подавлять HR, индуцированный Pseudomonas syringae pv. Phaseolus в растениях (Robinette and Matthysse, 1990). Это подавление зависит от активности онкогенов iaaH и iaaM , которые кодируют ферменты синтеза ауксина, поскольку несколько мутантов транспозона Agrobacterium в генах iaa не смогли ингибировать HR.Сходным образом, транскрипция нескольких генов, участвующих в передаче сигналов окислительного стресса, индуцируется только онкогенным, но не истощенным по Т-ДНК штаммом Agrobacterium (Lee et al., 2009). Производство H 2 O 2 предшествует HR, который расщепляется с помощью кодируемой хромосомой каталазы Agrobacterium (Xu and Pan, 2000). H 2 O 2 действует как локальный триггер запрограммированной гибели клеток и как распространяемый сигнал для индукции клеточных защитных генов в окружающих клетках (Levine et al., 1994). Помимо своих сигнальных функций, H 2 O 2 также участвует в укреплении клеточных стенок на начальных стадиях защиты растений путем перекрестного связывания структурных белков клеточной стенки (Bradley et al., 1992). Накопление H 2 O 2 предотвращается только на ранних стадиях агробактериальной инфекции, но продолжается по мере развития опухоли (Lee et al., 2009).
H 2 O 2 действует как локальный триггер запрограммированной гибели клеток и как распространяемый сигнал для индукции клеточных защитных генов в окружающих клетках (Levine et al., 1994). Помимо своих сигнальных функций, H 2 O 2 также участвует в укреплении клеточных стенок на начальных стадиях защиты растений путем перекрестного связывания структурных белков клеточной стенки (Bradley et al., 1992). Накопление H 2 O 2 предотвращается только на ранних стадиях агробактериальной инфекции, но продолжается по мере развития опухоли (Lee et al., 2009).
Ответы хоста на разработку Crown Gall
Морфологические адаптации
Развитие корончатых галлов сопровождается глубокими изменениями профиля экспрессии генов, метаболизма и морфологии.Неконтролируемый синтез ауксина и цитокинина клетками, трансформированными Т-ДНК онкогенных Ti-плазмид, стимулирует развитие опухоли, в то время как соотношение ауксина и цитокинина определяет морфологию корончатого галла (рис. 1В). В первые дни исследований молекулярных основ развития корончатых галлов было замечено, что мутации в локусе tmr , кодирующем ipt , вызывают корневые галлы, а мутации в локусах tms , кодирующих iaaH и iaaM вызывают фенотипы побегов (Garfinkel et al., 1981; Акиоши и др., 1983; Барри и др., 1984; Бухманн и др., 1985; Блэк и др., 1994). Недавнее исследование локуса Т-ДНК Atu6002 штамма C58 показало, что когда кодирующий белок C экспрессируется, он повышает чувствительность клетки-хозяина к ауксину (Lacroix et al., 2013). Помимо генов, кодируемых Т-ДНК, экспрессия нескольких генов-хозяев, участвующих в метаболизме и передаче сигналов ауксина и цитокинина, экспрессируется в корончатых галлах (Lee et al., 2009). Известно, что цитокинин и ауксин вместе с ET необходимы для роста опухолей коронарных галлов и дифференцировки типов клеток с различной морфологией и функциями (рис. 1C).В частности, было показано, что ЭТ необходим для формирования сосудистой ткани и развития опухоли корончатого желчного пузыря (Aloni et al.
1В). В первые дни исследований молекулярных основ развития корончатых галлов было замечено, что мутации в локусе tmr , кодирующем ipt , вызывают корневые галлы, а мутации в локусах tms , кодирующих iaaH и iaaM вызывают фенотипы побегов (Garfinkel et al., 1981; Акиоши и др., 1983; Барри и др., 1984; Бухманн и др., 1985; Блэк и др., 1994). Недавнее исследование локуса Т-ДНК Atu6002 штамма C58 показало, что когда кодирующий белок C экспрессируется, он повышает чувствительность клетки-хозяина к ауксину (Lacroix et al., 2013). Помимо генов, кодируемых Т-ДНК, экспрессия нескольких генов-хозяев, участвующих в метаболизме и передаче сигналов ауксина и цитокинина, экспрессируется в корончатых галлах (Lee et al., 2009). Известно, что цитокинин и ауксин вместе с ET необходимы для роста опухолей коронарных галлов и дифференцировки типов клеток с различной морфологией и функциями (рис. 1C).В частности, было показано, что ЭТ необходим для формирования сосудистой ткани и развития опухоли корончатого желчного пузыря (Aloni et al. , 1998; Wächter et al., 1999, 2003; Ullrich and Aloni, 2000). Применение ингибитора синтеза ET аминоэтоксивинил-глицина предотвращает васкуляризацию стеблей клещевины ( Ricinus communis ) и полностью подавляет рост опухоли (Wächter et al., 2003). Когда мутантный мутант Lycospersicon esculentum томата ( Lycospersicon esculentum ), нечувствительный к ET, инфицирован вирулентными клетками Agrobacterium , у него не развиваются опухоли, несмотря на интеграцию и экспрессию кодируемых Т-ДНК онкогенов для биосинтеза ауксина и цитокинина (Aloni и другие., 1998). Таким образом, неоваскуляризация является предпосылкой для развития корончатого галла.
, 1998; Wächter et al., 1999, 2003; Ullrich and Aloni, 2000). Применение ингибитора синтеза ET аминоэтоксивинил-глицина предотвращает васкуляризацию стеблей клещевины ( Ricinus communis ) и полностью подавляет рост опухоли (Wächter et al., 2003). Когда мутантный мутант Lycospersicon esculentum томата ( Lycospersicon esculentum ), нечувствительный к ET, инфицирован вирулентными клетками Agrobacterium , у него не развиваются опухоли, несмотря на интеграцию и экспрессию кодируемых Т-ДНК онкогенов для биосинтеза ауксина и цитокинина (Aloni и другие., 1998). Таким образом, неоваскуляризация является предпосылкой для развития корончатого галла.
Рост и распространение опухолей корончатого галла вызывают разрушение слоя эпидермальных клеток и, таким образом, потерю замыкающих клеток и интактной кутикулы. Соответственно экспрессия генов, участвующих в биосинтезе кутина, подавляется (Deeken et al., 2006). Поскольку нарушенная поверхность обеспечивает доступ патогенам и приводит к неконтролируемой потере воды растением-хозяином, поверхность корончатого галла необходимо герметизировать. Это достигается за счет дифференциации перидермоподобного поверхностного слоя (Ефетова и соавт., 2007). В полимеризации мономеров суберина участвуют пероксидазы, для которых H 2 O 2 является донором электронов. Таким образом, H 2 O 2 , продуцируемый в корончатых галлах, действует скорее на укрепление клеточных стенок, чем на индукцию HR. Стимулом для индуцирования суберизации является передача сигналов ABA, опосредованная засухой (рис. 1D). Сигнализация стресса от засухи, по-видимому, играет центральную роль в развитии корончатого галла. АБК накапливается в корончатых галлах в больших количествах, а транскрипция набора генов, индуцируемых засухой и/или АБК, повышена (Mistrik et al., 2000; Веселов и др., 2003; Ефетова и др., 2007). Синтез АБК запускается ЕТ, что продемонстрировано применением различных ингибиторов биосинтеза ЕТ или АБК, а также использованием нечувствительных к ЕТ или дефицитных по АБК мутантов томата (Hansen and Grossmann, 2000).
Это достигается за счет дифференциации перидермоподобного поверхностного слоя (Ефетова и соавт., 2007). В полимеризации мономеров суберина участвуют пероксидазы, для которых H 2 O 2 является донором электронов. Таким образом, H 2 O 2 , продуцируемый в корончатых галлах, действует скорее на укрепление клеточных стенок, чем на индукцию HR. Стимулом для индуцирования суберизации является передача сигналов ABA, опосредованная засухой (рис. 1D). Сигнализация стресса от засухи, по-видимому, играет центральную роль в развитии корончатого галла. АБК накапливается в корончатых галлах в больших количествах, а транскрипция набора генов, индуцируемых засухой и/или АБК, повышена (Mistrik et al., 2000; Веселов и др., 2003; Ефетова и др., 2007). Синтез АБК запускается ЕТ, что продемонстрировано применением различных ингибиторов биосинтеза ЕТ или АБК, а также использованием нечувствительных к ЕТ или дефицитных по АБК мутантов томата (Hansen and Grossmann, 2000). Среди генов, которые играют роль в защите опухолей корончатого галла от засухи, есть FAD3, , кодирующий десатуразу жирных кислот. У мутанта fad3-2 с нарушением биосинтеза α-линоленовой кислоты (C18:3) развиваются опухоли корончатого галла гораздо меньшего размера, особенно при низкой, но не высокой относительной влажности (Klinkenberg et al., 2014). Повышенные уровни C18:3 были обнаружены во фракции фосфолипидов опухолей корончатого галла Arabidopsis и сохраняют целостность мембраны в условиях засушливого стресса. В дополнение к изменениям экспрессии генов корончатые галлы накапливают большое количество осмопротекторов, таких как пролин (Pro), гамма-аминомасляная кислота (ГАМК) и альфа-аминоадипиновая кислота. Замедленный рост опухоли у мутантных растений abi и aba подчеркивает важность АБК-опосредованного сигнального пути стресса засухи в развитии корончатого галла (Ефетова и др., 2007).
Среди генов, которые играют роль в защите опухолей корончатого галла от засухи, есть FAD3, , кодирующий десатуразу жирных кислот. У мутанта fad3-2 с нарушением биосинтеза α-линоленовой кислоты (C18:3) развиваются опухоли корончатого галла гораздо меньшего размера, особенно при низкой, но не высокой относительной влажности (Klinkenberg et al., 2014). Повышенные уровни C18:3 были обнаружены во фракции фосфолипидов опухолей корончатого галла Arabidopsis и сохраняют целостность мембраны в условиях засушливого стресса. В дополнение к изменениям экспрессии генов корончатые галлы накапливают большое количество осмопротекторов, таких как пролин (Pro), гамма-аминомасляная кислота (ГАМК) и альфа-аминоадипиновая кислота. Замедленный рост опухоли у мутантных растений abi и aba подчеркивает важность АБК-опосредованного сигнального пути стресса засухи в развитии корончатого галла (Ефетова и др., 2007).
Перемещение питательных веществ и метаболизм
Профили экспрессии генов, участвующих в энергетическом метаболизме, таких как фотосинтез, митохондриальный транспорт электронов и ферментация, вместе с физиологическими данными показали, что опухоли Arabidopsis продуцируют соединения C и N гетеротрофно и получают энергию в основном анаэробно путем спиртового брожения (рис. 1E; Deeken et al. др., 2006). Переход от аутотрофии к гетеротрофии снижает уровень кислорода в опухолях корончатого галла, тем самым индуцируя экспрессию генов, чувствительных к гипоксии, таких как SAD6 .Этот ген кодирует стеароил-ацил-белок-носитель-десатуразу, которая принадлежит к классу ферментов, которые, как известно, катализируют первую стадию десатурации жирных кислот, кислородзависимый процесс. Несмотря на ограниченную доступность кислорода в корончатых галлах, SAD6 обеспечивает мононенасыщенную жирную кислоту, олеиновую кислоту, для мембранных фосфолипидов (Klinkenberg et al., 2014). Таким образом, экспрессия SAD6 поддерживает десатурацию жирных кислот в условиях гипоксии.
1E; Deeken et al. др., 2006). Переход от аутотрофии к гетеротрофии снижает уровень кислорода в опухолях корончатого галла, тем самым индуцируя экспрессию генов, чувствительных к гипоксии, таких как SAD6 .Этот ген кодирует стеароил-ацил-белок-носитель-десатуразу, которая принадлежит к классу ферментов, которые, как известно, катализируют первую стадию десатурации жирных кислот, кислородзависимый процесс. Несмотря на ограниченную доступность кислорода в корончатых галлах, SAD6 обеспечивает мононенасыщенную жирную кислоту, олеиновую кислоту, для мембранных фосфолипидов (Klinkenberg et al., 2014). Таким образом, экспрессия SAD6 поддерживает десатурацию жирных кислот в условиях гипоксии.
Опухоли корончатого галла в основном используют органический углерод и азот для роста и поэтому являются сильным поглотителем для растения-хозяина.Метаболиты и минералы должны быть предоставлены растением-хозяином и перемещены в опухоль коронного галла (рис. 1F). Механизмы переноса питательных веществ и их накопления были изучены на опухолях корончатого галла с применением цитологического окрашивания, электрофизиологических и 14 CO 2 методов трассировки, а также белка переноса вируса (Marz and Ullrich-Eberius, 1988; Malsy et al. и др., 1992; Прадель и др., 1999). Растворенные вещества попадают в опухоль корончатого галла через сосудистую ткань, которая связана с тканью растения-хозяина и состоит из флоэмы для транспорта ассимилятов и ксилемы для воды и минералов (Aloni et al., 1995; Дикен и др., 2003). Ассимиляты продуцируются исходными листьями и апопластически и симпластически выгружаются из флоэмы в опухолях корончатого галла. Высокая активность апопластной инвертазы указывает на то, что сахароза выгружается апопластически (Malsy et al., 1992). После расщепления сахарозы ферментами, расщепляющими сахарозу, гексозы могут быть поглощены переносчиками гексоз в опухолевые клетки. Корончатые галлы Arabidopsis демонстрируют повышенную экспрессию нескольких генов, кодирующих ферменты, разлагающие сахарозу, и переносчик моносахаридов (Deeken et al., 2006). Кроме того, высокоаффинный переносчик гексозы был выделен из меристематических клеток табака, трансформированных опухолью, индуцирующей Т-ДНК, и был охарактеризован как энергонезависимый переносчик поглощения гексозы (Verstappen et al.
и др., 1992; Прадель и др., 1999). Растворенные вещества попадают в опухоль корончатого галла через сосудистую ткань, которая связана с тканью растения-хозяина и состоит из флоэмы для транспорта ассимилятов и ксилемы для воды и минералов (Aloni et al., 1995; Дикен и др., 2003). Ассимиляты продуцируются исходными листьями и апопластически и симпластически выгружаются из флоэмы в опухолях корончатого галла. Высокая активность апопластной инвертазы указывает на то, что сахароза выгружается апопластически (Malsy et al., 1992). После расщепления сахарозы ферментами, расщепляющими сахарозу, гексозы могут быть поглощены переносчиками гексоз в опухолевые клетки. Корончатые галлы Arabidopsis демонстрируют повышенную экспрессию нескольких генов, кодирующих ферменты, разлагающие сахарозу, и переносчик моносахаридов (Deeken et al., 2006). Кроме того, высокоаффинный переносчик гексозы был выделен из меристематических клеток табака, трансформированных опухолью, индуцирующей Т-ДНК, и был охарактеризован как энергонезависимый переносчик поглощения гексозы (Verstappen et al. , 1991). Применение непроницаемого для мембраны флуоресцентного зонда, карбоксифлуоресцеина (CF) на исходных листьях и транзиторная экспрессия GFP-меченого белка оболочки (CP) вируса картофеля X (PVX), использующего исключительно плазмодесмы для распространения, продемонстрировали существование симпластного пути транспорта между флоэма и опухолевые клетки (Pradel et al., 1999). Оба репортера показывают обширное межклеточное движение в паренхиме опухолей корончатого галла, но не в неинфицированных тканях стебля различных видов растений, начиная от симпластических ( Curcubita maxima ) до апопластных загрузчиков ( R. communis, Nicotiana benthamiana ). Разрушенная и увеличенная поверхность опухоли корончатого галла приводит к перемещению воды и минералов в опухоли корончатого галла, поскольку скорость испарения корончатого галла превышает скорость испарения листьев и неинфицированных стеблей (Schurr et al., 1996; Вехтер и др., 2003). Похожий на перидерму слой опробковевших клеток, покрывающий поверхность корончатого галла, обеспечивает значительное диффузионное сопротивление водяному пару, но не является непроницаемым барьером для воды (рис.
, 1991). Применение непроницаемого для мембраны флуоресцентного зонда, карбоксифлуоресцеина (CF) на исходных листьях и транзиторная экспрессия GFP-меченого белка оболочки (CP) вируса картофеля X (PVX), использующего исключительно плазмодесмы для распространения, продемонстрировали существование симпластного пути транспорта между флоэма и опухолевые клетки (Pradel et al., 1999). Оба репортера показывают обширное межклеточное движение в паренхиме опухолей корончатого галла, но не в неинфицированных тканях стебля различных видов растений, начиная от симпластических ( Curcubita maxima ) до апопластных загрузчиков ( R. communis, Nicotiana benthamiana ). Разрушенная и увеличенная поверхность опухоли корончатого галла приводит к перемещению воды и минералов в опухоли корончатого галла, поскольку скорость испарения корончатого галла превышает скорость испарения листьев и неинфицированных стеблей (Schurr et al., 1996; Вехтер и др., 2003). Похожий на перидерму слой опробковевших клеток, покрывающий поверхность корончатого галла, обеспечивает значительное диффузионное сопротивление водяному пару, но не является непроницаемым барьером для воды (рис. 1D; Kolattukudy, Dean, 1974; Vogt et al., 1983; Schreiber). и др., 2005). Катионы и анионы поглощаются опухолевыми клетками благодаря функционированию локализованных в мембране каналов и транспортеров, экспрессируемых в корончатом галле (Deeken et al., 2003). Мутанты калиевых каналов с нарушенным ростом корончатых галлов подчеркивают важность оптимального снабжения питательными веществами для роста.
1D; Kolattukudy, Dean, 1974; Vogt et al., 1983; Schreiber). и др., 2005). Катионы и анионы поглощаются опухолевыми клетками благодаря функционированию локализованных в мембране каналов и транспортеров, экспрессируемых в корончатом галле (Deeken et al., 2003). Мутанты калиевых каналов с нарушенным ростом корончатых галлов подчеркивают важность оптимального снабжения питательными веществами для роста.
Ответы защиты
Гамма-аминомасляная кислота и Pro не только служат осмопротекторами в процессах, связанных со стрессом, связанным с засухой, у растения-хозяина, но также влияют на вирулентность Agrobacterium (Haudecoeur et al., 2009a, b). ГАМК, продуцируемый в опухолях корончатого галла, может поглощаться клетками Agrobacterium и вызывает задержку накопления 3-оксо-октаноилгомосеринлактона (OC8HSL) и конъюгации плазмиды Ti. ГАМК активирует оперон AttKLM, в котором лактоназа AttM разрушает сигнал восприятия кворума, OC8HSL, тем самым включая подавление кворума для защиты растения-хозяина от инфекций бактериальными патогенами (Yuan et al. , 2008). Однако Pro препятствует импорту ГАМК и тем самым предотвращает вызванную ГАМК деградацию сигнала восприятия бактериального кворума OC8HSL. Таким образом, Pro противодействует ГАМК-индуцированной деградации OC8HSL и, следовательно, может использоваться патогеном для обхода основанной на ГАМК защиты растения-хозяина.
, 2008). Однако Pro препятствует импорту ГАМК и тем самым предотвращает вызванную ГАМК деградацию сигнала восприятия бактериального кворума OC8HSL. Таким образом, Pro противодействует ГАМК-индуцированной деградации OC8HSL и, следовательно, может использоваться патогеном для обхода основанной на ГАМК защиты растения-хозяина.
В дополнение к процессам роста и развития, регулируемым ауксином и цитокинином, биология корончатых галлов также включает сигнальные пути защиты от патогенов. Гормоны, такие как SA, JA и ET, являются основными сигналами, вызывающими защитные реакции (López et al., 2008). У корончатых галлов Arabidopsis уровни SA и ET, но не JA, повышены (рис. 1G). JA не оказывает очевидного влияния на развитие опухоли корончатого галла, так как развитие Arabidopsis нечувствительных к JA мутантов сходно с диким типом (Lee et al., 2009). Содержание СА и ЭТ вместе с экспрессией патогенных маркерных генов СА- и ЭТ-зависимых сигнальных путей увеличивается по мере накопления транскриптов iaa и ipt, кодируемых Т-ДНК. Таким образом, ауксин и/или цитокинин, по-видимому, важны для передачи защитного сигнала в опухолях корончатого галла, поскольку неонкогенный штамм Agrobacterium , содержащий обезвреженный pTiC58, не индуцирует экспрессию маркерных генов SA- и ET-зависимых сигнальных путей. (Ли и др., 2009). Известно, что высокие уровни ауксина и цитокинина стимулируют синтез ЭТ и его накопление в корончатых галлах (Goodman et al., 1986; Aloni et al., 1998; Johnson, Ecker, 1998; Vogel et al., 1998; Wächter et al. ., 1999). В отличие от ЭТ классические маркерные гены СА-зависимых сигнальных путей не индуцируются, скорее всего, в результате высокого содержания ауксина, который, как было показано, ингибирует ответы СА, чтобы избежать индукции САР (Robert-Seilaniantz et al. ., 2011). Несмотря на отсутствие индукции SA-зависимой защитной сигнализации, мутантные растения Arabidopsis с высоким уровнем SA сильно снижают, в то время как растения с низким уровнем SA способствуют росту опухоли (Lee et al.
Таким образом, ауксин и/или цитокинин, по-видимому, важны для передачи защитного сигнала в опухолях корончатого галла, поскольку неонкогенный штамм Agrobacterium , содержащий обезвреженный pTiC58, не индуцирует экспрессию маркерных генов SA- и ET-зависимых сигнальных путей. (Ли и др., 2009). Известно, что высокие уровни ауксина и цитокинина стимулируют синтез ЭТ и его накопление в корончатых галлах (Goodman et al., 1986; Aloni et al., 1998; Johnson, Ecker, 1998; Vogel et al., 1998; Wächter et al. ., 1999). В отличие от ЭТ классические маркерные гены СА-зависимых сигнальных путей не индуцируются, скорее всего, в результате высокого содержания ауксина, который, как было показано, ингибирует ответы СА, чтобы избежать индукции САР (Robert-Seilaniantz et al. ., 2011). Несмотря на отсутствие индукции SA-зависимой защитной сигнализации, мутантные растения Arabidopsis с высоким уровнем SA сильно снижают, в то время как растения с низким уровнем SA способствуют росту опухоли (Lee et al. , 2009). Вместо того, чтобы индуцировать пути защиты хозяина, высокие уровни СА воздействуют непосредственно на онкогенные агробактерии, подавляя экспрессию гена vir и тем самым снижая вирулентность агробактерий (Yuan et al., 2007; Anand et al., 2008). Помимо опосредованного SA ингибирования вирулентности Agrobacterium , SA активирует оперон AttKLM, точно так же, как это делает ГАМК, для подавления чувства кворума у Agrobacterium (Yuan et al., 2008). Таким образом, активация подавления кворума ауксином, СК и ГАМК является частью программы защиты растений от Agrobacterium в развивающемся корончатом галле.В дополнение к SA, ET и IAA также ингибируют перенос регулона vir и Т-ДНК в растительные клетки (рис. 1G; Ezura et al., 2000; Nonaka et al., 2008). Таким образом, взаимодействие между растением-хозяином и Agrobacterium во многом основано на перекрестных помехах фитогормонов, которые обеспечивают баланс между защитой от патогенов хозяином и развитием корончатого галла, стимулируемым Agrobacterium .
, 2009). Вместо того, чтобы индуцировать пути защиты хозяина, высокие уровни СА воздействуют непосредственно на онкогенные агробактерии, подавляя экспрессию гена vir и тем самым снижая вирулентность агробактерий (Yuan et al., 2007; Anand et al., 2008). Помимо опосредованного SA ингибирования вирулентности Agrobacterium , SA активирует оперон AttKLM, точно так же, как это делает ГАМК, для подавления чувства кворума у Agrobacterium (Yuan et al., 2008). Таким образом, активация подавления кворума ауксином, СК и ГАМК является частью программы защиты растений от Agrobacterium в развивающемся корончатом галле.В дополнение к SA, ET и IAA также ингибируют перенос регулона vir и Т-ДНК в растительные клетки (рис. 1G; Ezura et al., 2000; Nonaka et al., 2008). Таким образом, взаимодействие между растением-хозяином и Agrobacterium во многом основано на перекрестных помехах фитогормонов, которые обеспечивают баланс между защитой от патогенов хозяином и развитием корончатого галла, стимулируемым Agrobacterium .
Эпигенетические процессы в интеграции ДНК, экспрессии онкогенов и развитии коронных галлов
Эпигенетические изменения, связанные с интеграцией Т-ДНК и экспрессией онкогена
Эпигенетические изменения, влияющие на структуру хроматина, играют важную роль в регуляции широкого спектра клеточных процессов.Например, гистоны подвергаются посттрансляционной модификации, включая ацетилирование, фосфорилирование, метилирование и убиквитинирование. Эти модификации могут влиять на развитие корончатых галлов на разных уровнях, либо влияя на структуру хроматина и интеграцию ДНК, либо влияя на экспрессию генов в ткани хозяина. Повышающая регуляция нескольких членов из семейств основных генов гистонов после заражения Agrobacterium указывает на то, что они важны для процесса трансформации (Veena et al., 2003). Например, мутантов Arabidopsis , лишенных гистона h3A, дефектны по интеграции Т-ДНК (Mysore et al., 2000). Кроме того, укороченная версия VIP1, белок Arabidopsis , предположительно взаимодействующий с Т-ДНК-белковым комплексом (Т-комплекс), который не способен взаимодействовать с гистоном h3A, сильно снижает туморогенность Agrobacterium (Li и др. , 2005). Поскольку это снижение, скорее всего, связано со снижением эффективности интеграции Т-ДНК, это предполагает, что ассоциация VIP1 с хроматином хозяина имеет решающее значение для интеграции Т-ДНК.Одна из гипотез того, как эпигенетическая информация влияет на интеграцию ДНК, заключается в том, что модификации хроматина, окружающие двухцепочечные разрывы (DSB) ДНК, могут быть распознаны Т-комплексом. Полученный хроматин-Т-комплекс может затем приблизить Т-ДНК к DSB и облегчить ее интеграцию путем репарации DSB (Magori and Citovsky, 2011). В качестве альтернативы гистоны могут также усиливать трансформацию, защищая поступающую ДНК от расщепления нуклеазами на начальных стадиях трансформации.В самом деле, избыточная экспрессия нескольких гистоновых генов в Arabidopsis приводит к увеличению количества перенесенной ДНК и повышенной транзиторной экспрессии трансгена в трансформированных клетках (Tenea et al., 2009). Другие эпигенетические модификации, такие как метилирование ДНК, не коррелируют с паттерном интеграции Т-ДНК, указывая на то, что интеграция Т-ДНК происходит независимо от этого типа модификации (Kim et al.
, 2005). Поскольку это снижение, скорее всего, связано со снижением эффективности интеграции Т-ДНК, это предполагает, что ассоциация VIP1 с хроматином хозяина имеет решающее значение для интеграции Т-ДНК.Одна из гипотез того, как эпигенетическая информация влияет на интеграцию ДНК, заключается в том, что модификации хроматина, окружающие двухцепочечные разрывы (DSB) ДНК, могут быть распознаны Т-комплексом. Полученный хроматин-Т-комплекс может затем приблизить Т-ДНК к DSB и облегчить ее интеграцию путем репарации DSB (Magori and Citovsky, 2011). В качестве альтернативы гистоны могут также усиливать трансформацию, защищая поступающую ДНК от расщепления нуклеазами на начальных стадиях трансформации.В самом деле, избыточная экспрессия нескольких гистоновых генов в Arabidopsis приводит к увеличению количества перенесенной ДНК и повышенной транзиторной экспрессии трансгена в трансформированных клетках (Tenea et al., 2009). Другие эпигенетические модификации, такие как метилирование ДНК, не коррелируют с паттерном интеграции Т-ДНК, указывая на то, что интеграция Т-ДНК происходит независимо от этого типа модификации (Kim et al. , 2007). Что касается посттрансляционных модификаций гистонов, РНК-опосредованный нокдаун двух гистоновых деацетилаз (HDT1 и HDT2) снижает эффективность опосредованной Agrobacterium трансформации корневых сегментов Arabidopsis (Crane and Gelvin, 2007).Деацетилирование гистонов функционирует при компактизации хроматина и репрессии транскрипции (Strahl and Allis, 2000). Следовательно, наблюдаемый эффект на трансформацию может быть результатом воздействия на структуру хроматина или экспрессии генов растительных факторов, участвующих в процессе интеграции. Деацетилирование гистонов также может влиять на интеграцию ДНК, воздействуя на репарацию DSB, поскольку несколько деацетилаз гистонов имеют решающее значение для процесса репарации ДНК у дрожжей (Munoz-Galvan et al., 2013).
, 2007). Что касается посттрансляционных модификаций гистонов, РНК-опосредованный нокдаун двух гистоновых деацетилаз (HDT1 и HDT2) снижает эффективность опосредованной Agrobacterium трансформации корневых сегментов Arabidopsis (Crane and Gelvin, 2007).Деацетилирование гистонов функционирует при компактизации хроматина и репрессии транскрипции (Strahl and Allis, 2000). Следовательно, наблюдаемый эффект на трансформацию может быть результатом воздействия на структуру хроматина или экспрессии генов растительных факторов, участвующих в процессе интеграции. Деацетилирование гистонов также может влиять на интеграцию ДНК, воздействуя на репарацию DSB, поскольку несколько деацетилаз гистонов имеют решающее значение для процесса репарации ДНК у дрожжей (Munoz-Galvan et al., 2013).
После того, как Т-ДНК интегрируется в геном растения, растение-хозяин часто подавляет трансгены.Замалчивание генов может происходить по двум различным механизмам. Сайленсинг генов транскрипции (TGS) является результатом инактивации промотора, в то время как пост-TGS (PTGS) происходит, когда промотор активен, но мРНК не накапливается. Метилирование ДНК промоторных последовательностей часто связано с инактивацией трансгенов (Linne et al., 1990; Matzke and Matzke, 1991; Kilby et al., 1992). Скрининг большой коллекции трансгенных линий Arabidopsis с одиночными копиями Т-ДНК, включая репортерный ген pNOS- NPTII , показал, что метилирование промотора необходимо, но недостаточно для инактивации транскрипции (Fischer et al., 2008). Замалчивание происходит только тогда, когда растения, зараженные трансгеном-сайленсером, также обеспечивают сигнал РНК. Что касается локальных особенностей генома хозяина, влияющих на замалчивание генов, повторы, фланкирующие место интеграции, по-видимому, способствуют инактивации, тогда как фланкирующие гены скорее ее ослабляют. Сайленсинг РНК запускается только в том случае, если уровень транскрипта трансгена превышает специфический для гена порог, что позволяет предположить, что инактивация является частью механизма защиты растений, соответствующего чрезмерно транскрибируемым генам (Schubert et al.
Метилирование ДНК промоторных последовательностей часто связано с инактивацией трансгенов (Linne et al., 1990; Matzke and Matzke, 1991; Kilby et al., 1992). Скрининг большой коллекции трансгенных линий Arabidopsis с одиночными копиями Т-ДНК, включая репортерный ген pNOS- NPTII , показал, что метилирование промотора необходимо, но недостаточно для инактивации транскрипции (Fischer et al., 2008). Замалчивание происходит только тогда, когда растения, зараженные трансгеном-сайленсером, также обеспечивают сигнал РНК. Что касается локальных особенностей генома хозяина, влияющих на замалчивание генов, повторы, фланкирующие место интеграции, по-видимому, способствуют инактивации, тогда как фланкирующие гены скорее ее ослабляют. Сайленсинг РНК запускается только в том случае, если уровень транскрипта трансгена превышает специфический для гена порог, что позволяет предположить, что инактивация является частью механизма защиты растений, соответствующего чрезмерно транскрибируемым генам (Schubert et al. , 2004).
, 2004).
Помимо подавляющей регуляции трансгенов, интегрированных в геном растения вместе с Т-ДНК, сама Т-ДНК также может подвергаться модификации механизмом молчания растений. Первый комплексный анализ метилирования Т-ДНК показал, что метилирование может происходить в различных линиях опухолей растений, индуцированных Agrobacterium . По крайней мере, одна копия Т-ДНК в геноме каждой опухоли оставалась неметилированной, что позволяло экспрессировать онкоген и пролиферировать корончатые галлы (Gelvin et al., 1983). Эксперименты с использованием деметилирующего агента 5-азацитидина показывают, что метилирование отрицательно коррелирует с экспрессией генов в опухолях растений (Hepburn et al., 1983). Более недавнее исследование метилирования Т-ДНК в опухолях корончатых галлов, индуцированных на стеблях Arabidopsis , демонстрирует, что последовательности онкогенов метилированы только до очень низкой степени (Gohlke et al., 2013). Две межгенные области, которые служат промоторами для экспрессии онкогенов iaaH , iaaM, и ipt, , полностью неметилированы в корончатых галлах Arabidopsis . Поскольку генные продукты этих онкогенов необходимы для повышения уровней цитокинина и ауксина, они всегда активно транскрибируются в опухолях корончатых галлов стеблей Arabidopsis (Deeken et al., 2006). Низкая степень метилирования Т-ДНК в корончатых галлах позволяет предположить, что это является предпосылкой для поддержания уровней экспрессии онкогенов, необходимых для образования опухолей. Действительно, индукции метилирования ДНК онкогена путем продукции двухцепочечных РНК достаточно, чтобы подавить транскрипцию онкогена и предотвратить развитие опухоли (Gohlke et al., 2013).
Поскольку генные продукты этих онкогенов необходимы для повышения уровней цитокинина и ауксина, они всегда активно транскрибируются в опухолях корончатых галлов стеблей Arabidopsis (Deeken et al., 2006). Низкая степень метилирования Т-ДНК в корончатых галлах позволяет предположить, что это является предпосылкой для поддержания уровней экспрессии онкогенов, необходимых для образования опухолей. Действительно, индукции метилирования ДНК онкогена путем продукции двухцепочечных РНК достаточно, чтобы подавить транскрипцию онкогена и предотвратить развитие опухоли (Gohlke et al., 2013).
Эпигенетические модификации в геноме коронарных галлов
Анализ стеблей соцветий, инфицированных Agrobacterium , позволил отслеживать экспрессию генов в опухоли корончатого галла на более поздних стадиях развития и выявил массивные изменения в ее транскриптоме (Deeken et al., 2006). Было обнаружено, что большая часть генома Arabidopsis (около 22% генов) по-разному экспрессируется между корончатыми галлами и ложно инфицированными стеблями. Было обнаружено, что из этих генов несколько более высокий процент подавляется в корончатых галлах (12%) по сравнению с генами с повышающей регуляцией (10%).Отчетливые изменения экспрессии происходят в генах, имеющих ключевое значение для энергетического метаболизма, таких как участвующие в фотосинтезе, митохондриальном переносе электронов и ферментации. Это отражает индуцированные клетками-хозяевами изменения от ауксотрофного, аэробного метаболизма к гетеротрофному, транспортно-зависимому, сахарозависимому анаэробному метаболизму (см. Перемещение питательных веществ и метаболизм).
Было обнаружено, что из этих генов несколько более высокий процент подавляется в корончатых галлах (12%) по сравнению с генами с повышающей регуляцией (10%).Отчетливые изменения экспрессии происходят в генах, имеющих ключевое значение для энергетического метаболизма, таких как участвующие в фотосинтезе, митохондриальном переносе электронов и ферментации. Это отражает индуцированные клетками-хозяевами изменения от ауксотрофного, аэробного метаболизма к гетеротрофному, транспортно-зависимому, сахарозависимому анаэробному метаболизму (см. Перемещение питательных веществ и метаболизм).
Учитывая, что высокий процент генома Arabidopsis по-разному регулируется в опухолях корончатых галлов, перепрограммирование транскрипции, вероятно, происходит на нескольких уровнях.Например, уровни транскриптов нескольких семейств факторов транскрипции (MYB, bHLH, bZIP, домен AP) изменяются после инфицирования Agrobacterium (Ditt et al., 2006; Sardesai et al., 2013), тем самым индуцируя опухолеспецифический ген. образец выражения. Экспрессия генов также может регулироваться эпигенетическими механизмами, такими как модификация хроматина или метилирование ДНК. Помимо модификаций, которые играют роль во время интеграции Т-ДНК и подавления онкогенов (см. Эпигенетические процессы в интеграции ДНК, экспрессии онкогенов и развитии коронных галлов), метилирование ДНК генов растений также может влиять на рост опухоли (рис. 1H).Действительно, 8% генов, кодирующих белок, по-разному метилированы в корончатых галлах по сравнению с ложно инфицированными стеблями с общей тенденцией к гиперметилированию (Gohlke et al., 2013). В зависимости от положения метилирования ДНК наблюдаются разные эффекты на уровни экспрессии генов. В соответствии с тенденциями, наблюдаемыми для изменений метилирования ДНК в Arabidopsis (Zhang et al., 2006), повышенное метилирование в сайтах начала и конца транскрипции оказывает негативное влияние на экспрессию генов, в то время как эти два процесса положительно коррелируют в транскрибируемой области.
образец выражения. Экспрессия генов также может регулироваться эпигенетическими механизмами, такими как модификация хроматина или метилирование ДНК. Помимо модификаций, которые играют роль во время интеграции Т-ДНК и подавления онкогенов (см. Эпигенетические процессы в интеграции ДНК, экспрессии онкогенов и развитии коронных галлов), метилирование ДНК генов растений также может влиять на рост опухоли (рис. 1H).Действительно, 8% генов, кодирующих белок, по-разному метилированы в корончатых галлах по сравнению с ложно инфицированными стеблями с общей тенденцией к гиперметилированию (Gohlke et al., 2013). В зависимости от положения метилирования ДНК наблюдаются разные эффекты на уровни экспрессии генов. В соответствии с тенденциями, наблюдаемыми для изменений метилирования ДНК в Arabidopsis (Zhang et al., 2006), повышенное метилирование в сайтах начала и конца транскрипции оказывает негативное влияние на экспрессию генов, в то время как эти два процесса положительно коррелируют в транскрибируемой области. Картирование метилирования ДНК в опухолях выявило гипометилирование в восходящих областях генов, а также гиперметилирование в транскрибируемых областях. Оба они могут, в свою очередь, влиять на экспрессию генов и вносить свой вклад в опухолеспецифический паттерн экспрессии. Неудивительно, что пути, которые связаны с развитием опухоли, такие как гены, связанные с клеточным делением, биотическим стрессом и окислительно-восстановительной регуляцией, по-разному метилированы. Изменения в характере метилирования также влияют на рост опухоли, поскольку мутанты Arabidopsis в путях метилирования de novo способствуют развитию корончатых галлов.Интересно, что индукция каллуса, которая, как и развитие корончатого галла, также связана с дедифференцировкой растительных клеток, увеличивается у мутанта метилтрансферазы cmt3 (Berdasco et al., 2008). Кроме того, обработка ингибитором метилтрансферазы 5-акацитидином приводит к повышенному образованию мозолей. Недавно паттерн метилирования ДНК был тщательно изучен в каллусе Populus trichocarpa и Oryza sativa .
Картирование метилирования ДНК в опухолях выявило гипометилирование в восходящих областях генов, а также гиперметилирование в транскрибируемых областях. Оба они могут, в свою очередь, влиять на экспрессию генов и вносить свой вклад в опухолеспецифический паттерн экспрессии. Неудивительно, что пути, которые связаны с развитием опухоли, такие как гены, связанные с клеточным делением, биотическим стрессом и окислительно-восстановительной регуляцией, по-разному метилированы. Изменения в характере метилирования также влияют на рост опухоли, поскольку мутанты Arabidopsis в путях метилирования de novo способствуют развитию корончатых галлов.Интересно, что индукция каллуса, которая, как и развитие корончатого галла, также связана с дедифференцировкой растительных клеток, увеличивается у мутанта метилтрансферазы cmt3 (Berdasco et al., 2008). Кроме того, обработка ингибитором метилтрансферазы 5-акацитидином приводит к повышенному образованию мозолей. Недавно паттерн метилирования ДНК был тщательно изучен в каллусе Populus trichocarpa и Oryza sativa . У каллусов Oryza sativa было обнаружено гиперметилирование по сравнению с растениями дикого типа (Stroud et al., 2013). Генные тела гиперметилированы в каллусе Populus trichocarpa по сравнению с эксплантами, в то время как метилирование промотора снижено (Vining et al., 2013). В соответствии с паттерном метилирования корончатых галлов, гиперметилирование ДНК, по-видимому, является общим признаком дедифференцированного статуса.
У каллусов Oryza sativa было обнаружено гиперметилирование по сравнению с растениями дикого типа (Stroud et al., 2013). Генные тела гиперметилированы в каллусе Populus trichocarpa по сравнению с эксплантами, в то время как метилирование промотора снижено (Vining et al., 2013). В соответствии с паттерном метилирования корончатых галлов, гиперметилирование ДНК, по-видимому, является общим признаком дедифференцированного статуса.
Попытка идентифицировать внутренние сигналы растений, которые могут влиять на метилирование ДНК, предполагает, что высокие уровни АБК вызывают метилирование ДНК промоторных последовательностей (рис. 1H; Gohlke et al., 2013). Следовательно, этот фитогормон может быть, по крайней мере, частично ответственным за характер метилирования, обнаруживаемый в корончатых галлах. Заманчиво предположить, что АБК индуцирует метилирование ДНК в ответ на абиотические стрессы, такие как акклиматизация к засухе из-за повышенной потери воды в опухолях корончатого галла (Schurr et al. , 1996). Возможно, сигнальные пути АБК взаимосвязаны с процессами метилирования в корончатых галлах, как предполагалось для Physcomitrella patens (Khraiwesh et al., 2010). В будущем было бы интересно проанализировать мутанты с нокаутом ABA в отношении их паттерна метилирования, чтобы всесторонне картировать изменения метилирования, вызванные ABA, и тем самым улучшить наше понимание связи между различными путями. Кроме того, было бы интересно изучить влияние других фитогормонов на характер метилирования ДНК у корончатых галлов, поскольку они обнаруживают повышенный уровень не только АБК, но и цитокинина, ауксина, ЭТ и ЖК (Веселов и др., 2003; Ли и др., 2009).
, 1996). Возможно, сигнальные пути АБК взаимосвязаны с процессами метилирования в корончатых галлах, как предполагалось для Physcomitrella patens (Khraiwesh et al., 2010). В будущем было бы интересно проанализировать мутанты с нокаутом ABA в отношении их паттерна метилирования, чтобы всесторонне картировать изменения метилирования, вызванные ABA, и тем самым улучшить наше понимание связи между различными путями. Кроме того, было бы интересно изучить влияние других фитогормонов на характер метилирования ДНК у корончатых галлов, поскольку они обнаруживают повышенный уровень не только АБК, но и цитокинина, ауксина, ЭТ и ЖК (Веселов и др., 2003; Ли и др., 2009).
Сводка и Outlook
В начале заражения обнаружение Agrobacterium не вызывает сильной защитной реакции растения-хозяина. Agrobacterium скорее использует защитные реакции для повышения восприимчивости хозяина к трансформации и сигнальные пути хозяина для стимулирования роста бактерий. Однако у корончатых галлов пути защиты от патогенов значительно активируются и подавляют вирулентность Agrobacterium . Соответственно, растение-хозяин способно ограничивать количество дальнейших событий трансформации Т-ДНК и контролировать размеры роста корончатых галлов, которые представляют собой сильный метаболический сток для растения-хозяина. Метаболические и морфологические адаптации сопровождают развитие корончатых галлов и создают ткань, ориентированную на импорт. Гетеротрофный метаболизм вместе с анаэробным получением энергии требует транслокации метаболитов, воды и минералов из растения в пролиферирующую ткань корончатого галла.В качестве основы для перемещения питательных веществ сосудистая ткань должна дифференцироваться, а разрушенная и опробкованная поверхность корончатого галла обеспечивает движущую силу для потока питательных веществ. Фактически, опробкованная поверхность сводит к минимуму потери воды, но при этом позволяет достаточное количество воды испаряться. Целостность мембран поддерживается при низком уровне кислорода и повышенном уровне АФК в корончатых галлах за счет адаптации метаболизма липидов.
Соответственно, растение-хозяин способно ограничивать количество дальнейших событий трансформации Т-ДНК и контролировать размеры роста корончатых галлов, которые представляют собой сильный метаболический сток для растения-хозяина. Метаболические и морфологические адаптации сопровождают развитие корончатых галлов и создают ткань, ориентированную на импорт. Гетеротрофный метаболизм вместе с анаэробным получением энергии требует транслокации метаболитов, воды и минералов из растения в пролиферирующую ткань корончатого галла.В качестве основы для перемещения питательных веществ сосудистая ткань должна дифференцироваться, а разрушенная и опробкованная поверхность корончатого галла обеспечивает движущую силу для потока питательных веществ. Фактически, опробкованная поверхность сводит к минимуму потери воды, но при этом позволяет достаточное количество воды испаряться. Целостность мембран поддерживается при низком уровне кислорода и повышенном уровне АФК в корончатых галлах за счет адаптации метаболизма липидов. Изменения транскрипции, лежащие в основе физиологических изменений, частично вызваны дифференциальным метилированием ДНК генома корончатой галлы.В заключение, как инфекция Agrobacterium , так и рост корончатого галла являются строго регулируемыми процессами, которые сопровождаются защитой хозяина от патогенов и контрзащитой, запускаемой Agrobacterium . Эта регуляция происходит на разных уровнях, включая эпигенетический контроль экспрессии генов, изменения содержания фитогормонов, а также метаболические и морфологические адаптации.
Изменения транскрипции, лежащие в основе физиологических изменений, частично вызваны дифференциальным метилированием ДНК генома корончатой галлы.В заключение, как инфекция Agrobacterium , так и рост корончатого галла являются строго регулируемыми процессами, которые сопровождаются защитой хозяина от патогенов и контрзащитой, запускаемой Agrobacterium . Эта регуляция происходит на разных уровнях, включая эпигенетический контроль экспрессии генов, изменения содержания фитогормонов, а также метаболические и морфологические адаптации.
Несмотря на то, что Agrobacterium -растение-взаимодействие изучается более 100 лет и, скорее всего, является одним из самых известных взаимодействий патоген-хозяин, все еще остается несколько вопросов, на которые можно попытаться ответить. .В дополнение к вопросу о роли фитогормонов, отличных от АБК, в метилировании ДНК в развитии корончатого галла, еще один вопрос касается молекулярных механизмов того, как клетки Agrobacterium продуцируют ауксин и как ауксин повышает восприимчивость хозяина к трансформации. Более того, статус и тип растительных клеток, восприимчивых к интеграции Т-ДНК, пока неизвестны. Знание о клеточной идентичности, чувствительной к трансформации, улучшит наше понимание устойчивых к трансформации видов растений.Более того, процессы дифференцировки в корончатых галлах не следуют обычному паттернированию, в отличие от ситуации в органах растений, где паттернирование развития лежит в основе точной пространственно-временной экспрессии сигналов и родственных им рецепторов. Поскольку первоначальная/типичная программа развития, по-видимому, отвергается, опухоли корончатого галла предоставляют уникальную возможность для изучения молекулярных и биохимических механизмов, лежащих в основе клеточной дедифференцировки, а также процессов дифференцировки. Не все поднятые вопросы могут быть легко разрешены, поскольку некоторые из них требуют сложных методов, которые сначала необходимо разработать и внедрить.Однако изобретение новых методов принесет пользу всему научному сообществу, как это было раньше, когда Agrobacterium стал биотехнологическим инструментом для создания генетически модифицированных растений.
Более того, статус и тип растительных клеток, восприимчивых к интеграции Т-ДНК, пока неизвестны. Знание о клеточной идентичности, чувствительной к трансформации, улучшит наше понимание устойчивых к трансформации видов растений.Более того, процессы дифференцировки в корончатых галлах не следуют обычному паттернированию, в отличие от ситуации в органах растений, где паттернирование развития лежит в основе точной пространственно-временной экспрессии сигналов и родственных им рецепторов. Поскольку первоначальная/типичная программа развития, по-видимому, отвергается, опухоли корончатого галла предоставляют уникальную возможность для изучения молекулярных и биохимических механизмов, лежащих в основе клеточной дедифференцировки, а также процессов дифференцировки. Не все поднятые вопросы могут быть легко разрешены, поскольку некоторые из них требуют сложных методов, которые сначала необходимо разработать и внедрить.Однако изобретение новых методов принесет пользу всему научному сообществу, как это было раньше, когда Agrobacterium стал биотехнологическим инструментом для создания генетически модифицированных растений.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Исследования проводились при финансовой поддержке Deutsche Forschungsgemeinschaft (проект B5, SFB 567 и проект A5 Исследовательской учебной группы 1342 — Передача сигналов липидов).Мы благодарим Стэнтона Гелвина за критическое прочтение рукописи.
Каталожные номера
Акиоши Д.Э., Моррис Р., Хинц Р., Мишке Б., Косуге Т., Гарфинкель Д. и соавт. (1983). Цитокинин/ауксиновый баланс в опухолях корончатого галла регулируется специфическими локусами в Т-ДНК. Проц. Натл. акад. науч. США 80, 407–411. doi: 10.1073/pnas.80.2.407
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Акиёси, Д. Э., Регьер, Д. А., и Гордон, М.П. (1987). Продукция цитокинина Agrobacterium и Pseudomonas spp. J. Бактериол. 169, 4242–4248.
J. Бактериол. 169, 4242–4248.
Опубликовано Резюме | Опубликован полный текст
Акиёси Д.Э., Регьер Д.А., Джен Г. и Гордон М.П. (1985). Клонирование и нуклеотидная последовательность гена tzs из штамма Agrobacterium tumefaciens T37. Рез. нуклеиновых кислот. 13, 2773–2788. дои: 10.1093/нар/13.8.2773
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Алони, Р., Pradel, K.S., and Ullrich, C.I. (1995). Трехмерная структура сосудистых тканей в Agrobacterium tumefaciens , индуцированных корончатых галлах, и в стеблях-хозяевах Ricinus communis L. Planta 196, 597–605. дои: 10.1007/BF00203661
Полнотекстовая перекрестная ссылка
Алони Р., Вольф А., Фейгенбаум П., Авни А. и Клее Х. Дж. (1998). Никогда не созревший мутант свидетельствует о том, что этилен, индуцированный опухолью, контролирует морфогенез индуцированных Agrobacterium tumefaciens корончатых галлов на стеблях томатов. Завод физиол. 117, 841–849. doi: 10.1104/стр.117.3.841
Завод физиол. 117, 841–849. doi: 10.1104/стр.117.3.841
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ананд, А., Уппалапати, С.Р., Рю, С.-М., Аллен, С.Н., Канг, Л., Тан, Ю., и др. (2008). Салициловая кислота и системная приобретенная резистентность играют роль в ослаблении болезни корончатого галла, вызываемой Agrobacterium tumefaciens . Завод физиол. 146, 703–715. doi: 10.1104/стр.107.111302
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Барри, Г.Ф., Роджерс С.Г., Фрэйли Р.Т. и Бранд Л. (1984). Идентификация клонированного гена биосинтеза цитокинина. Проц. Натл. акад. науч. США 81, 4776–4780. doi: 10.1073/pnas.81.15.4776
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Бердаско М., Алькасар Р., Гарсия-Ортис М.В., Баллестар Э., Фернандес А.Ф., Ролдан-Архона Т. и др. (2008). Гиперметилирование промоторной ДНК и репрессия генов в недифференцированных клетках Arabidopsis . PLoS ONE 3:e3306. doi: 10.1371/journal.pone.0003306
PLoS ONE 3:e3306. doi: 10.1371/journal.pone.0003306
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Биннс, А. Н., и Костантино, П. (1998). «Онкогены Agrobacterium », в The Rhizobiaceae , под ред. Х. П. Спаник, А. Кондороси и П. Дж. Дж. Хойкаас (Нидерланды: Springer), 251–266.
Black, R.C., Binns, A.N., Chang, C.F., and Lynn, D.G. (1994). Клеточно-автономный цитокинин-независимый рост клеток табака, трансформированных штаммом Agrobacterium tumefaciens , лишенным гена биосинтеза цитокинина. Завод физиол. 105, 989–998. doi: 10.1104/стр.105.3.989
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Брэдли, Д. Дж., Кьеллбом, П., и Лэмб, С. Дж. (1992). Вызываемое элиситором и раной окислительное сшивание богатого пролином белка клеточной стенки растений: новая быстрая защитная реакция. Сотовый 70, 21–30. дои: 10.1016/0092-8674(92)
-PОпубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Бухманн И., Марнер Ф.Дж., Шредер Г., Ваффеншмидт С. и Шредер Дж. (1985). Гены опухолей у растений: Т-ДНК кодирует биосинтез цитокинина. EMBO J. 4, 853–859.
Опубликовано Резюме | Опубликован полный текст
Шато С., Сангван Р. С. и Сангван-Норрел Б. С. (2000). Компетентность генотипов и мутантов Arabidopsis thaliana в отношении опосредованного переноса генов Agrobacterium tumefaciens : роль фитогормонов. Дж. Эксп. Бот. 51, 1961–1968. дои: 10.1093/jexbot/51.353.1961
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Chilton, M.D., Drummond, M.H., Merio, D.J., Sciaky, D., Montoya, A.L., Gordon, M.P., et al. (1977). Стабильное включение плазмидной ДНК в клетки высших растений: молекулярная основа онкогенеза корончатых галлов. Сотовый 11, 263–271. дои: 10.1016/0092-8674(77)
-5
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Шиншилла Д., Бауэр З., Регенасс М., Боллер Т.и Феликс Г. (2006). Рецепторная киназа Arabidopsis FLS2 связывает flg22 и определяет специфичность восприятия флагеллина. Растительная клетка 18, 465–476. doi: 10.1105/tpc.105.036574
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Крейн, Ю.М., и Гелвин, С. (2007). РНКи-опосредованное замалчивание генов выявляет участие генов, связанных с хроматином Arabidopsis , в опосредованной Agrobacterium корневой трансформации. Проц.Натл. акад. науч. США 104. doi: 10.1073/pnas.0706986104
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Deeken, R., Engelmann, J.C., Efetova, M., Czirjak, T., Müller, T., Kaiser, W.M., et al. (2006). Интегрированное представление об экспрессии генов и профилях растворенных веществ в опухолях Arabidopsis : полногеномный подход. Растительная клетка 18, 3617–3634. doi: 10.1105/tpc.106.044743
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Дикен, Р., Ивашикина Н., Циряк Т., Филиппар К., Беккер Д., Аче П. и соавт. (2003). В развитии опухоли Arabidopsis thaliana участвуют Shaker-подобные K+ каналы AKT1 и AKT2/3. Завод J. 34, 778–787. doi: 10.1046/j.1365-313X.2003.01766.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Дитт Р.Ф., Керр К.Ф., де Фигейредо П., Делроу Дж., Комай Л. и Нестер Э.В. (2006). Транскриптом Arabidopsis thaliana в ответ на Agrobacterium tumefaciens . Мол. Взаимодействие растительных микробов. 19, 665–681. doi: 10.1094/MPMI-19-0665
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Джамей, А., Питчке, А., Накагами, Х., Радж, И., и Хирт, Х. (2007). Стратегия троянского коня в трансформации Agrobacterium : злоупотребление защитной сигнализацией MAPK. Наука 318, 453–456. doi: 10.1126/наука.1148110
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ефетова М., Zeier, J., Riederer, M., Lee, C.-W., Stingl, N., Mueller, M., et al. (2007). Центральная роль абсцизовой кислоты в защите от стресса засухи Agrobacterium -индуцированных опухолей на Arabidopsis . Завод физиол. 145, 853–862. doi: 10.1104/стр.107.104851
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Эзура Х., Юхаси К.-И., Ясута Т. и Минамисава К. (2000). Влияние этилена на перенос генов, опосредованный Agrobacterium tumefaciens , в дыню. Порода растений. 119, 75–79. doi: 10.1046/j.1439-0523.2000.00438.x
Полнотекстовая перекрестная ссылка
Феликс Г., Дюран Дж. Д., Волко С. и Боллер Т. (1999). Растения имеют чувствительную систему восприятия наиболее консервативного домена бактериального флагеллина. Завод J. 18, 265–276. doi: 10.1046/j.1365-313X.1999.00265.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Фишер У., Кульманн М., Печинка А., Шмидт Р. и Метте М.Ф. (2008). Локальные особенности ДНК влияют на РНК-направленное подавление транскрипционных генов и метилирование ДНК. Завод Ж. 53, 1–10. doi: 10.1111/j.1365-313X.2007.03311.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Гафни Ю., Ихт М. и Рубинфельд Б.-З. (1995). Стимуляция вирулентности Agrobacterium tumefaciens индол-3-уксусной кислотой. Письмо. заявл. микробиол. 20, 98–101. doi: 10.1111/j.1472-765X.1995.tb01295.x
Полнотекстовая перекрестная ссылка
Гарфинкель, Д.Дж., Симпсон Р.Б., Рим Л.В., Уайт Ф.Ф., Гордон М.П. и Нестер Э.В. (1981). Генетический анализ корончатого галла: карта тонкой структуры Т-ДНК с помощью сайт-направленного мутагенеза. сотовый 27, 143–153. дои: 10.1016/0092-8674(81)
-8Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Гельвин, С. Б. (2000). Agrobacterium и гены растений, участвующие в переносе и интеграции Т-ДНК. Анну. Преподобный Завод Физиол. Завод Мол. биол. 51, 223–256.doi: 10.1146/annurev.arplant.51.1.223
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Гелвин, С., Карчер, С.Дж., и ДиРита, В.Дж. (1983). Метилирование Т-ДНК в Agrobacterium tumefaciens и в нескольких опухолях корончатого галла. Рез. нуклеиновых кислот. 11, 159–174. doi: 10.1093/нар/11.1.159
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Гольке, Дж., Шольц, К.-Дж., Кнейц, С., Вебер, Д., Фукс, Дж., Хедрих, Р., и другие. (2013). Опосредованный метилированием ДНК контроль экспрессии генов имеет решающее значение для развития опухолей корончатого галла. Генетика PLoS. 9:e1003267. doi: 10.1371/journal.pgen.1003267
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Гомес-Гомес, Л., Бауэр, З., и Боллер, Т. (2001). И внеклеточный домен повторов, богатый лейцином, и киназная активность FSL2 необходимы для связывания флагеллина и передачи сигналов у Arabidopsis . Растительная клетка 13, 1155–1163.
Гомес-Гомес Л., Феликс Г. и Боллер Т. (1999). Единственный локус определяет чувствительность к бактериальному флагеллину у Arabidopsis thaliana . Завод J. 18, 277–284. doi: 10.1046/j.1365-313X.1999.00451.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Гудман Т.С., Монтойя А.Л., Уильямс С. и Чилтон М.Д. (1986). Устойчивая продукция этилена в дисках моркови, трансформированных Agrobacterium , вызвана экспрессией продуктов гена Т-ДНК tms. J. Бактериол. 167, 387–388.
Опубликовано Резюме | Опубликован полный текст
Грей, Дж., Гелвин, С.Б., Мейлан, Р., и Моррис, Р.О. (1996). Транспортная РНК является источником внеклеточного изопентениладенина в Ti-бесплазмидном штамме Agrobacterium tumefaciens . Завод физиол. 110, 431–438. doi: 10.1104/стр.110.2.431
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Одекер, Э., Планаменте, С., Сиру, А., Таньер, М., Shelp, B.J., Moréra, S., et al. (2009а). Пролин противодействует индуцированному ГАМК подавлению чувства кворума у Agrobacterium tumefaciens . Проц. Натл. акад. науч. США 106, 14587–14592. doi: 10.1073/pnas.0808005106
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Haudecoeur, E., Tannières, M., Cirou, A., Raffoux, A., Dessaux, Y., and Faure, D. (2009b). Различная регуляция и роль лактоназ AiiB и AttM в Agrobacterium tumefaciens C58. Мол. Взаимодействие растительных микробов. 22, 529–537. doi: 10.1094/MPMI-22-5-0529
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Хепберн, А.Г., Кларк, Л.Е., Пирсон, Л., и Уайт, Дж. (1983). Роль метилирования цитозина в контроле экспрессии гена нопалинсинтазы в опухоли растений. Дж. Мол. заявл. Жене. 2, 315–329.
Опубликовано Резюме | Опубликован полный текст
Хван, Х.-Х., Ван, М.-Х., Ли, Ю.-Л., Цай, Ю.-Л., Ли, Ю.-Х., Ян, Ф.-Дж. и др. (2010). Agrobacterium — продуцируемые и экзогенно-модулированные цитокинином Agrobacterium — опосредованная трансформация растений. Мол. Завод Патол. 11, 677–690. doi: 10.1111/j.1364-3703.2010.00637.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Хван, Х.-Х., Ян, Ф.-Дж., Ченг, Т.-Ф., Чен, Ю.-К., Ли, Ю.-Л., Цай, Ю.-Л., и др. др. (2013). Белок Tzs и экзогенный цитокинин влияют на экспрессию генов вирулентности и рост бактерий Agrobacterium tumefaciens . Фитопатология 103, 888–899. doi: 10.1094/PHYTO-01-13-0020-R
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Janssens, A., Engler, G., Zambryski, P., and Montagu, M.V. (1984). Область Т-ДНК нопалина С58 транскрибируется в Agrobacterium tumefaciens . Мол. Генерал Жене. 195, 341–350. дои: 10.1007/BF00332769
Полнотекстовая перекрестная ссылка
Храйвеш Б., Ариф М. А., Сеумел Г. И., Оссовски С., Вейгель Д., Рески Р. и соавт. (2010). Транскрипционный контроль экспрессии генов с помощью микроРНК. Сотовый 140, 111–122. doi: 10.1016/j.cell.2009.12.023
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ким С.-И., Вина и Гелвин С.Б. (2007). Полногеномный анализ сайтов интеграции Т-ДНК Agrobacterium в геноме Arabidopsis , созданном в неселективных условиях. Завод J. 51, 779–791. doi: 10.1111/j.1365-313X.2007.03183.х
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Клинкенберг Дж., Фаист Х., Сауп С., Ламбертс С., Кришке М., Стингл Н. и др. (2014). Две десатуразы жирных кислот, SAD6 и FAD3, участвуют в передаче сигналов стресса от засухи и гипоксии у корончатых галлов Arabidopsis . Завод физиол. 164, 570–583. doi: 10.1104/стр.113.230326
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Колаттукуди, П. Э., и Дин, Б.Б. (1974). Структура, газохроматографическое измерение и функция суберина, синтезируемого ломтиками ткани клубня картофеля. Завод физиол. 54, 116–121. doi: 10.1104/pp.54.1.116
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Кренс Ф.А., Трифонова А., Пол Кейзер Л.К. и Холл Р.Д. (1996). Влияние экзогенно применяемых фитогормонов на эффективность переноса генов у сахарной свеклы ( Beta vulgaris L.). Растениевод. 116, 97–106.дои: 10.1016/0168-9452(96)04357-9
Полнотекстовая перекрестная ссылка
Кулдау, Г. А., Де Вос, Г., Оуэн, Дж., Маккефри, Г., и Замбриски, П. (1990). Оперон virB Agrobacterium tumefaciens pTiC58 кодирует 11 открытых рамок считывания. Мол. Генерал Жене. МГГ 221, 256–266. дои: 10.1007/BF00261729
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Кунце Г., Ципфель К., Робацек С., Нихаус К., Боллер Т. и Феликс Г. (2004).N-конец бактериального фактора элонгации Tu вызывает врожденный иммунитет у растений Arabidopsis . Растительная клетка 16, 3496–3507. doi: 10.1105/tpc.104.026765
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Кутаек М. и Ровенска Дж. (1991). Синтез ауксина в трансформированной растительной ткани Agrobacterium tumefaciens и A. tumefaciens. Регулятор роста растений. 10, 313–327. дои: 10.1007/BF00024591
Полнотекстовая перекрестная ссылка
Лакруа, Б., Гизатуллина Д.И., Бабст Б.А., Гиффорд А.Н., Цитовский В. (2013). Agrobacterium Белок Atu6002, кодируемый Т-ДНК, препятствует ответу хозяина на ауксин. Мол. Завод Патол. 15, 275–283. doi: 10.1111/mpp.12088
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Lee, C.-W., Efetova, M., Engelmann, J.C., Kramell, R., Wasternack, C., Ludwig-Muller, J., et al. (2009). Agrobacterium tumefaciens способствует индукции опухоли путем модулирования защиты от патогенов у Arabidopsis thaliana . Растительная клетка 21, 2948–2962. doi: 10.1105/tpc.108.064576
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Левин А., Тенхакен Р., Диксон Р. и Лэмб К. (1994). H 2 O 2 от окислительного взрыва управляет реакцией растения на гиперчувствительность к болезням. Сотовый 79, 583–593. дои: 10.1016/0092-8674(94)
-4Полнотекстовая перекрестная ссылка
Ли Дж., Кричевский А., Вайдья М., Цфира Т. и Читовский В.(2005). Разобщение функций белка Arabidopsis VIP1 при транзиторной и стабильной генетической трансформации растений с помощью Agrobacterium . Проц. Натл. акад. науч. США 102, 5733–5738. doi: 10.1073/pnas.0404118102
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Линн Ф., Хайдманн И., Сэдлер Х. и Мейер П. (1990). Эпигенетические изменения в экспрессии гена кукурузы А1 у петунии гибридной: роль числа интегрированных копий гена и степень метилирования. Мол. Генерал Жене. МГГ 222, 329–336. дои: 10.1007/BF00633837
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Лю П. и Нестер Э. В. (2006). Индолуксусная кислота, продукт перенесенной ДНК, ингибирует экспрессию гена vir и рост Agrobacterium tumefaciens C58. Проц. Натл. акад. науч. США 103, 4658–4662. doi: 10.1073/pnas.0600366103
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Лю, С.Т. и Кадо, С.И. (1979). Производство индолуксусной кислоты: функция плазмиды Agrobacterium tumefaciens C58. Биохим. Биофиз. Рез. коммун. 90, 171–178. дои: 10.1016/0006-291X(79)-X
Полнотекстовая перекрестная ссылка
Лопес, М. А., Банненберг, Г., и Кастресана, К. (2008). Контроль передачи сигналов гормонов является проблемой роста и выживания растений и патогенов. Курс. мнение биол. растений 11, 420–427. doi: 10.1016/j.pbi.2008.05.002
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Магори, С.и Цитовски В. (2012). Роль убиквитин-протеасомной системы в Agrobacterium tumefaciens -опосредованной генетической трансформации растений. Завод физиол. 160, 65–71. doi: 10.1104/pp.112.200949
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Мальси С., Бел В., Клюге М., Хартунг В. и Ульрих С.И. (1992). Индукция корончатых галлов Agrobacterium tumefaciens (штамм C58) обращает транслокацию и накопление ассимилятов в Kalanchoë daigremontiana . Окружающая среда растительных клеток. 15, 519–529. doi: 10.1111/j.1365-3040.1992.tb01485.x
Полнотекстовая перекрестная ссылка
Марц, С., и Ульрих-Эбериус, К.И. (1988). Накопление растворенных веществ и электрический мембранный потенциал в корончатых галлах, индуцированных Agrobacterium tumefaciens , в листьях Kalanchoë daigremontiana . Растениевод. 57, 27–36. дои: 10.1016/0168-9452(88)-0
Полнотекстовая перекрестная ссылка
Мацке, М. А., и Мацке, А. Дж. М.(1991). Дифференциальная инактивация и метилирование трансгена в растениях двумя супрессорными локусами, содержащими гомологичные последовательности. Завод Мол. биол. 16, 821–830. дои: 10.1007/BF00015074
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Мистрик И., Павловкин Дж., Вехтер Р., Прадель К.С., Швалм К., Хартунг В. и соавт. (2000). Влияние стволовых опухолей, индуцированных Agrobacterium tumefaciens , на поглощение NO 3 — у Ricinus communis . Почва для растений 226, 87–98. дои: 10.1023/A:1026465606865
Полнотекстовая перекрестная ссылка
Моррис, Р. О. (1986). Гены, определяющие биосинтез ауксинов и цитокининов у фитопатогенов. Анну. Преподобный Завод Физиол. 37, 509–538. doi: 10.1146/annurev.pp.37.060186.002453
Полнотекстовая перекрестная ссылка
Муньос-Гальван С., Химено С., Ротштейн Р. и Агилера А. (2013). Ацетилирование гистона h4K56, Rad52 и факторы репарации, не относящиеся к ДНК, контролируют выбор репарации двухцепочечных разрывов с помощью сестринской хроматиды. Генетика PLoS. 9:e1003237. doi: 10.1371/journal.pgen.1003237
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Майсур, К.С., Нам, Дж., и Гелвин, С.Б. (2000). Мутант гистона h3A Arabidopsis дефектен в интеграции Т-ДНК Agrobacterium . Проц. Натл. акад. науч. США 97, 948–953. doi: 10.1073/pnas.97.2.948
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Нонака С., Юхаши К.-И., Такада К., Сугаваре М., Минамисава К. и Эзура Х. (2008). Продукция этилена в растениях во время трансформации подавляет экспрессию гена vir у Agrobacterium tumefaciens . Новый Фитол. 178, 647–656. doi: 10.1111/j.1469-8137.2008.02400.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Pradel, K.S., Ullrich, C.I., Cruz, S.S., and Oparka, K.J. (1999). Симпластическая непрерывность в опухолях, индуцированных Agrobacterium tumefaciens . Дж. Эксп. Бот. 50, 183–192. doi: 10.1093/jxb/50.331.183
Полнотекстовая перекрестная ссылка
Роберт-Сейланианц, А., Грант, М., и Джонс, Дж. Д. Г. (2011). Гормональные перекрестные помехи при заболеваниях и защите растений: больше, чем просто антагонизм жасмоната и салицилата. Анну. Преподобный Фитопат. 49, 317–343. doi: 10.1146/annurev-phyto-073009–114447
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Робинетт, Д., и Маттис, А. Г. (1990). Ингибирование Agrobacterium tumefaciens и Pseudomonas savastanoi развития реакции гиперчувствительности, вызываемой Pseudomonas syringae pv. фазоликола . J. Бактериол. 172, 5742–5749.
Опубликовано Резюме | Опубликован полный текст
Сардесай, Н., Ли, Л.-Ю., Чен, Х., Йи, Х., Ольбрихт, Г.Р., Штирнберг, А., и соавт. (2013). Цитокинины, секретируемые Agrobacterium , способствуют трансформации путем репрессии растительного фактора транскрипции Myb. Науч. Сигнал. 6, ра100. doi: 10.1126/scisignal.2004518
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Шрайбер, Л., Франке Р. и Хартманн К. (2005). Восковое и субериновое развитие нативной и раневой перидермы картофеля ( Solanum tuberosum L.) и его связь с перидермальной транспирацией. Планта 220, 520–530. doi: 10.1007/s00425-004-1364-9
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Шредер, Г., Клипп, В., Хиллебранд, А., Эринг, Р., Конц, С. и Шредер, Дж. (1983). Консервативная часть Т-области Ti-плазмид у бактерий экспрессирует четыре белка. EMBO J. 2, 403–409.
Шуберт Д., Лехтенберг Б., Форсбах А., Гилс М., Бахадур С. и Шмидт Р. (2004). Сайленсинг в трансформантах Т-ДНК Arabidopsis – преобладающая роль ген-специфического механизма восприятия РНК по сравнению с эффектами положения. Растительная клетка 104.024547. doi: 10.1105/tpc.104.024547
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Шурр У., Шуберт Б., Алони Р., Прадель К. С., Шмундт Д., Jaehne, B., et al. (1996). Структурные и функциональные доказательства ксилем-опосредованного транспорта воды и высокой транспирации в Agrobacterium tumefaciens -индуцированных опухолях Ricinus communis . Бот. Acta 109, 405–411. doi: 10.1111/j.1438-8677.1996.tb00590.x
Полнотекстовая перекрестная ссылка
Ширасу, К., Морел, П., и Кадо, К.И. (1990). Характеристика оперона virB плазмиды Agrobacterium tumefaciens Ti: последовательность нуклеотидов и анализ белков. Мол. микробиол. 4, 1153–1163. doi: 10.1111/j.1365-2958.1990.tb00690.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Стаскавич, Б.Дж., Осубель, Ф.М., Бейкер, Б.Дж., Эллис, Дж.Г., и Джонс, Дж.Д. (1995). Молекулярная генетика устойчивости растений к болезням. Наука 268, 661–667. doi: 10.1126/science.7732374
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Степанова А.Н., Хойт Дж.М., Гамильтон А.А.и Алонсо, Дж. М. (2005). Связь между этиленом и ауксином обнаружена при характеристике двух специфичных для корней мутантов, нечувствительных к этилену, у Arabidopsis . Растительная клетка 17, 2230–2242. doi: 10.1105/tpc.105.033365
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Страуд, Х., Дин, Б., Саймон, С.А., Фенг, С., Беллицци, М., Пеллегрини, М., и др. (2013). Растения-регенеранты из культуры тканей риса содержат стабильные эпигеномные изменения. eLife 2:e00354. doi: 10.7554/eLife.00354
Опубликовано Резюме | Опубликован полный текст
Tenea, G.N., Spantzel, J., Lee, L.-Y., Zhu, Y., Lin, K., Johnson, S.J., et al. (2009). Сверхэкспрессия нескольких генов гистонов Arabidopsis увеличивает опосредованную Agrobacterium трансформацию и экспрессию трансгена в растениях. Растительная клетка 21, 3350–3367. doi: 10.1105/tpc.109.070607
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Томасшоу, М.Ф., Хагли С., Бухгольц В.Г. и Томасшоу Л.С. (1986). Молекулярная основа ауксин-независимого фенотипа опухолевых тканей корончатого галла. Наука 231, 616–618. doi: 10.1126/science.3511528
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Томпсон Д.В., Мелчерс Л.С., Идлер К.Б., Шильпероорт Р.А. и Хойкаас П.Дж. (1988). Анализ полной нуклеотидной последовательности оперона Agrobacterium tumefaciens virB. Рез. нуклеиновых кислот. 16, 4621–4636. doi: 10.1093/нар/16.10.4621
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Veena, Jiang, H., Doerge, R.W., and Gelvin, S.B. (2003). Перенос Т-ДНК и белков Vir в растительные клетки с помощью Agrobacterium tumefaciens индуцирует экспрессию генов-хозяев, участвующих в опосредовании трансформации, и подавляет экспрессию генов защиты хозяина. Завод J. 35, 219–236. doi: 10.1046/j.1365-313X.2003.01796.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ферстаппен, Р., Раностай С. и Рауш Т. (1991). Транспортеры гексозы на плазматической мембране и тонопласте трансформированных растительных клеток: кинетическая характеристика двух разных переносчиков. Биохим. Биофиз. Acta 1073, 366–373. дои: 10.1016/0304-4165(91)-6
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Веселов, Д., Лангханс, М., Хартунг, В., Алони, Р., Фойснер, И., Гётц, К., и соавт. (2003). Развитие Agrobacterium tumefaciens C58-индуцированных опухолей растений и воздействие на побеги-хозяева контролируются каскадом жасмоновой кислоты, ауксина, цитокинина, этилена и абсцизовой кислоты. Планта 216, 512–522. doi: 10.1007/s00425-002-0883-5
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Vining, K., Pomraning, K.R., Wilhelm, L.J., Ma, C., Pellegrini, M., Di, Y., et al. (2013). Реорганизация метилома при дедифференцировке и регенерации in vitro Populus trichocarpa . BMC Растение Биол. 13:92. дои: 10.1186/1471-2229-13-92
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Фогель, Дж.П., Шуерман П., Вёсте К., Брандстаттер И. и Кибер Дж. Дж. (1998). Выделение и характеристика мутантов Arabidopsis , дефектных по индукции биосинтеза этилена цитокинином. Генетика 149, 417–427.
Опубликовано Резюме | Опубликован полный текст
Vogt, E., Schönherr, J., and Schmidt, H.W. (1983). Водопроницаемость мембран перидермы, выделенных ферментативным путем из клубней картофеля ( Solanum tuberosum L.). Планта 158, 294–301.дои: 10.1007/BF00397330
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Wächter, R., Fischer, K., Gäbler, R., Kühnemann, F., Urban, W., Bögemann, G.M., et al. (1999). Продукция этилена и накопление АСС в опухолях растений, индуцированных Agrobacterium tumefaciens , и их влияние на структуру и функцию опухоли и стебля хозяина. Окружающая среда растительных клеток. 22, 1263–1273. doi: 10.1046/j.1365-3040.1999.00488.x
Полнотекстовая перекрестная ссылка
Вехтер, Р., Langhans, M., Aloni, R., Götz, S., Weilmünster, A., Koops, A., et al. (2003). Васкуляризация, поток большого объема раствора и локальные роли ферментов метаболизма сахарозы во время онкогенеза с помощью Agrobacterium tumefaciens . Завод физиол. 133, 1024–1037. doi: 10.1104/стр.103.028142
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Уорд, Д. В., Дрейпер, О., Зупан, Дж. Р., и Замбрыски, П. К. (2002). Картирование пептидной связи системы секреции IV типа Agrobacterium tumefaciens vir, кодирующей vir, выявляет белковые субсборки. Проц. Натл. акад. науч. США 99, 11493–11500. doi: 10.1073/pnas.1723
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ward, J.E., Akiyoshi, D.E., Regier, D., Datta, A., Gordon, M.P., and Nester, E.W. (1988). Характеристика оперона virB из плазмиды Agrobacterium tumefaciens Ti. Дж. Биол. хим. 263, 5804–5814.
Опубликовано Резюме | Опубликован полный текст
Вуд, Д. В., Сетубал, Дж. К., Kaul, R., Monks, D.E., Kitajima, J.P., Okura, V.K., et al. (2001). Геном природного генного инженера Agrobacterium tumefaciens C58. Наука 294, 2317–2323. doi: 10.1126/science.1066804
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Юань, З.-К., Эдлинд, М.П., Лю, П., Саенхам, П., Банта, Л.М., Уайз, А.А., и соавт. (2007). Сигнальная салициловая кислота растений подавляет экспрессию регулона vir и активирует гены подавления квормонов в Agrobacterium . Проц. Натл. акад. науч. США 104, 11790–11795. doi: 10.1073/pnas.0704866104
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Юань, З. К., Одекур, Э., Фор, Д., Керр, К. Ф., и Нестер, Э. В. (2008). Сравнительный транскриптомный анализ Agrobacterium tumefaciens в ответ на сигналы растения салициловой кислотой, индол-3-уксусной кислотой и гамма-аминомасляной кислотой выявляет сигнальные перекрестные помехи и коэволюцию Agrobacterium -растения. Клеточная микробиология. 10, 2339–2354. doi: 10.1111/j. 1462-5822.2008.01215
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Zhang, X., Yazaki, J., Sundaresan, A., Cokus, S., Chan, S.W.-L., Chen, H., et al. (2006). Полногеномное картирование с высоким разрешением и функциональный анализ метилирования ДНК у Arabidopsis . Сотовый 126, 1189–1201. doi: 10.1016/j.cell.2006.08.003
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ципфель, К., Kunze, G., Chinchilla, D., Caniard, A., Jones, J.D.G., Boller, T., et al. (2006). Восприятие бактериального PAMP EF-Tu рецептором EFR ограничивает Agrobacterium -опосредованную трансформацию. Сотовый 125, 749–760. doi: 10.1016/j.cell.2006.03.037
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Zipfel, C., Robatzek, S., Navarro, L., Oakeley, E.J., Jones, J.D.G., Felix, G., et al. (2004). Устойчивость к бактериальным заболеваниям у Arabidopsis за счет восприятия флагеллина. Природа 428, 764–767. doi: 10.1038/nature02485
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Плюсы Минусы, Побочные эффекты, Шрамы и Последующий уход
Кому следует избегать процедуры установки зубной коронки?
Лица должны избегать процедуры, если:
Заядлые курильщики; это может вызвать дегенерацию короны. В таких случаях металлическая коронка будет наиболее подходящей.
Не соблюдают гигиену полости рта; если не соблюдать гигиену полости рта, коронка может легко загрязниться.
Склонны к кариесу и подобным заболеваниям; если за коронкой образуется полость, в конечном итоге коронку, возможно, придется удалить.
У них нет крепких десен и зубов; слабые зубы или десны могут вызвать ненужное кровотечение и отек после установки коронки.
Не имеют или не могут избавиться от вредных привычек, таких как кусание ногтей, стискивание зубов и использование зубов для открывания упаковки.
Какие альтернативы зубной коронке?
Ниже приведены альтернативы, позволяющие избежать затрат времени и средств на получение коронки:
Пломбирование зубов
Если зуб сломан или разрушается, самое простое и дешевое – удалить дефектную часть и запломбировать полость.Это не долгосрочное решение и требует регулярной доработки. Пломбирование, как правило, делается при помощи коронки, но может выполняться отдельно, чтобы выиграть время, пока вы не получите более убедительное лечение.
Фарфоровые накладки/вкладки
Это направлено на сохранение большей части вашей естественной формы зубов. Накладки добавляются к разрушенной или сломанной части зуба, тогда как вкладки заполняют полые полости. Фарфор плотно прилегает к существующей зубной эмали. Эта техника требует высокого уровня навыков и является прекрасной альтернативой зубной коронке.
Временные коронки
Стоматолог также может предложить временные коронки, которые приклеиваются к зубам до тех пор, пока пациент не найдет окончательное решение. Это временные коронки, которые обычно держатся от 3 до 9 месяцев.
Удаление зуба
Если состояние зуба не поддается лечению, лучше его просто удалить. После удаления пустое пространство может быть занято имплантатами или зубными мостами или их комбинацией.
Каковы временные побочные эффекты зубных коронок?
После установки зубной коронки может возникнуть несколько распространенных осложнений.Вот некоторые из них:
Аллергическая реакция
Аллергическая реакция чаще всего вызвана материалом, из которого изготовлена коронка. В результате зуб плохо прилегает к коронке. В таком случае свяжитесь с врачом как можно скорее, чтобы определить подходящий курс действий.
Дискомфорт/чувствительность
Любой дискомфорт или чувствительность обычно возникают из-за присутствия инородного предмета во рту.Чтобы облегчить это, регулярно принимайте лекарства, назначенные врачом. Кроме того, в течение недели ешьте мягкую пищу, такую как супы и мороженое, чтобы облегчить процесс адаптации.
Свободная коронка
Избегайте употребления липкой пищи в дни после процедуры, так как это может привести к расшатыванию коронки до того, как она прочно приклеится к зубу. Если коронка расшатана, это может привести к дополнительным рискам, таким как образование полости между коронкой и зубом. Убедитесь, что вы сообщаете своему врачу о расшатанной коронке, как только вы это заметите, чтобы избежать какой-либо инфекции.
Сколотая коронка
Сколы коронки часто являются результатом плохо проведенной процедуры. Коронка сколется, если хирург не сможет аккуратно установить коронку. Иногда коронка также может сколоться, если вы слишком сильно жуете или кусаете. В таком случае замените коронку до того, как это приведет к образованию полости. Кроме того, убедитесь, что ваш хирург имеет необходимый опыт для процедуры.
Как выбрать хирурга для процедуры зубной коронки?
Выбор хирурга для зубной коронки не должен быть сложной задачей, будь то в вашей стране или в стране за границей.Процедура относительно проста и не требует от медицинского персонала большого опыта. Однако, выбирая хирурга за границей, убедитесь, что он соответствует международным стандартам лечения и ухода. Вы можете сделать это, проверив его / ее сертификацию в качестве практикующего врача, в дополнение к их образованию, а также статус аккредитации больницы на международной платформе. Вы даже можете пообщаться с врачом и получить представление о его/ее практических знаниях о процедуре.
С посредниками по медицинскому туризму выбрать хирурга так же просто, сидя за тысячи километров, как и лично встретившись с ним/ней.
Должна ли ванная комната иметь молдинг
Должен ли в ванной комнате быть молдинг короны
Можете ли вы сделать форму короны в ванной комнате?
Хотя ванные комнаты, как правило, намного меньше по размеру, добавление формы короны может обеспечить еще больший способ увеличить пространство.Это может быть подчеркивание верхнего края комнаты или скоса, но венчание также дает эффект значительного увеличения высоты потолка (что всегда является плюсом).
Стоит ли добавлять молдинги в этом отношении?
Чтобы ответить на вопрос просто, формирование кроны не является общим решением. Приятно иметь его в одних комнатах, не используя в других. Некоторые комнаты в доме почти всегда являются популярным местом для формирования кроны. Например, гостиная — классическое место для его использования.
Точно так же, где вы размещаете форму короны?
Как было сказано выше, молдинги часто устанавливаются между стеной и потолком или между стеной и полом. Это придает помещению однородный и элегантный вид.
Где должна быть установлена рама?
- Кухонная мебель.
- Зеркало в ванной.
- Изголовье спальни.
- Доски.
- Все на вашем смонтированном телевизоре.
Тогда можно также спросить: увеличивает ли образование кроны меновую стоимость?
Установите правильно — несмотря на то, что другие могут сказать вам, молдинги короны довольно сложны в установке.Профессиональная покраска. И последнее, но не менее важное: молдинги могут повысить ценность вашего дома только в том случае, если они профессионально окрашены. Плохой цвет ничего не сделает с вашим домом — или того хуже.
Кастинг короны в моде?
Если у вас литые детали коронки = литые детали коронки никогда не выйдут из моды. Единственная причина, по которой вы больше не можете этого видеть, заключается в том, что люди не могут себе этого позволить. Если у них нет формирования кроны = формирование кроны устарело. Правда = наверное зависит от стиля дома и качества изготовления.
Нужно ли красить форму короны перед установкой?
Окрашенные стены
Вам нужен формирователь, чтобы соответствовать плинтусам?
Нет, точное соответствие декоративной планки и плинтуса не требуется. Заливка и резка могут варьироваться от толстого до тонкого, от широкого до узкого. Регулировка размеров и стиля оформления карнизов и молдингов придает помещению приятный эстетический вид.
Какова цель формирования кроны?
Потолочные молдинги представляют собой декоративный отделочный элемент, обычно используемый для покрытия шкафов, колонн и, как правило, внутренних стен в местах, где стена встречается с потолком.Он используется только в верхней части комнаты, поэтому термин корона используется для описания украшения комнаты.
Делает ли корона комнату больше?
Коронация в большинстве случаев не уменьшает комнату. Молдинг может быть шире, если крыша немного выше, и может достигать 6 дюймов в ширину для очень высокой крыши до 12 футов. Широкие рамки подходят только для очень больших комнат или очень высоких потолков, или и того, и другого.
Имеют ли ремесленные домики форму кроны?
Какую форму короны мне следует использовать?
Полосы желоба обычно имеют ширину от 3 до 20 дюймов.Используйте высоту комнаты, чтобы определить ширину формы кроны. Установите форму для короны шириной от 3 до 5 3/4 дюймов в комнате с потолком высотой восемь футов.
Современный дом имеет форму короны?
Молдинги с глубокими пятнами распространены в традиционных домах. Однако, если у вас современный дом и вы хотите теплоты цветных деревянных гирлянд, использование очень тонкого плоского профиля придает дереву очень современный вид. Использование простых плоских или фигурных молдингов для современных интерьеров — не единственный вариант.
Должен ли я удалить форму короны?
Корона представляет собой обычный декоративный молдинг между верхом стены и потолком. При покраске или другом ремонте стен иногда необходимо удалить плесень. Удаление формы коронки без повреждения формы или стенки — трудоемкий процесс.
Что можно использовать вместо формы короны?
Другой популярной альтернативой традиционным трехрядным литым деталям является древесноволокнистая плита средней плотности (МДФ).Устанавливается как настоящий молдинг, но дешевле и легче. Молдинги короны из МДФ также очень индивидуальны, поскольку их можно тонировать или красить в любой цвет, который вы хотите.
Нужно ли отливать корону?
Сколько я должен заплатить, чтобы разместить форму короны?
Средняя стоимость установки пресс-формы для коронки составляет 7-16 за погонный фут, в зависимости от выбранного материала и стоимости рабочей силы. Для средней гостиной лепнина стоит от 464 до 1102, кухня — от 210 до 570, а весь дом — от 2000 до 4000 за работу и установку.
Форма короны все еще популярна?
Несмотря на то, что молдинги всегда популярны, избегайте их использования в комнатах этого типа, так как они могут сделать комнату слишком тесной.
Какова ценность молдингов в вашем доме?
Ориентировочная стоимость: 5 долларов США за погонный метр
Замена обивки дорого?
Средняя стоимость замены
Под каким углом вы срезаете форму кроны?
Как я могу вырезать и повесить форму короны?
Для установки формы коронки:
Кухня имеет форму короны?
Должна ли ванная комната иметь молдингкрон | Фонд здоровья полости рта
Что такое корона?Коронка – это искусственная реставрация, которая надевается на оставшуюся часть отпрепарированного зуба, делая его прочным и придавая ему форму натурального зуба.Корону иногда называют «кепкой».
Зачем мне корона?Коронки — идеальный способ восстановить зубы, которые были сломаны или ослаблены кариесом или очень большой пломбой. Корона могла использоваться по ряду других причин, например:
- У вас может измениться цвет пломбы, и вы хотите улучшить внешний вид зуба.
- Возможно, у вас была пломбировка корня, и вам нужна коронка, чтобы защитить то, что осталось от зуба.
- Это может помочь надежно удерживать мост или зубной протез на месте.
Коронки могут быть изготовлены из различных материалов, и постоянно появляются новые материалы. Некоторые из наиболее популярных вариантов перечислены ниже.
Фарфор, соединенный с драгоценным металлом:Из этого материала делают большинство коронок. Изготавливается основа из драгоценного металла, а затем на нее слоями наносится фарфор.
ФарфорЭти коронки полностью изготовлены из фарфора и не так прочны, как коронки с фиксацией. Но они могут выглядеть очень естественно и чаще всего используются для передних зубов.
ЦельнокерамическийЭта современная техника предлагает безметалловую альтернативу, которая обеспечивает прочность коронки с фиксацией и внешний вид фарфоровой коронки. Поэтому он подходит для использования во всех областях рта.
Стекло №Эти коронки выглядят очень естественно и могут использоваться в любой части рта.
Коронки из сплава золотаЗолото – один из старейших пломбировочных материалов. Сегодня он используется с другими металлическими сплавами для увеличения его прочности, что делает его очень износостойким. Эти короны серебряного или золотого цвета.
Как готовят коронку?Стоматолог подготовит зуб к идеальной форме коронки. Это будет включать удаление слоя внешней поверхности, оставляя прочное внутреннее ядро. Количество удаленного зуба будет таким же, как толщина коронки.
После того, как зуб будет сформирован, команда стоматологов сделает оттиск (слепок) препарированного зуба, одного из противоположной челюсти и, возможно, еще одного, чтобы показать, как вы прикусываете друг друга.
Затем слепки будут переданы зубному технику вместе с информацией об используемом оттенке и любой другой необходимой информацией.
Что такое почтовая коронка?В зубах с запломбированными корнями может потребоваться установка штифта в корень зуба перед установкой коронки.Стойка обеспечивает поддержку и помогает коронке оставаться на месте. Поверхность зуба может быть удалена до уровня десны.
Штифт может быть изготовлен из предварительно изготовленной нержавеющей стали, и стоматолог может установить его непосредственно в корневой канал. Кроме того, зубной техник может изготовить штифт по индивидуальному заказу, который точно соответствует форме подготовленного корневого канала. Штифт помещается в корневой канал и цементируется в положении, готовом к установке коронки.
Существуют ли альтернативы штифтовым коронкам для зубов с запломбированными корнями?Если запломбированный корень зуба не разрушен полностью, стоматолог может восстановить его заново, используя пломбировочный материал.Затем этот «стержень» подготавливается так же, как и натуральный зуб, и снимаются оттиски.
Что будет происходить между посещениями?Будет изготовлена временная коронка, чтобы вы могли пользоваться зубом в ожидании изготовления коронки. Эта корона может быть более заметной, но только временной.
Как устанавливается коронка?Когда вы и ваш стоматолог довольны тем, что новая коронка подходит и выглядит хорошо, она будет зафиксирована на месте с помощью специального стоматологического цемента или клея.Цемент образует уплотнение, удерживающее коронку на месте.
Сколько времени занимает лечение?Вам потребуется как минимум два визита. При первом посещении команда стоматологов подготовит зуб, снимет слепки, заметит оттенок вашего зуба и установит временную коронку. Во время второго визита стоматолог установит постоянную коронку. Между визитами обычно проходит 1-2 недели.
Больно ли препарировать зуб под коронку?№У вас будет местная анестезия, и подготовительная работа не должна отличаться от пломбирования. Если в зубе нет нерва, а готовится штифтовая коронка, то местная анестезия может и не понадобиться.
Будет ли заметна корона?Коронка будет максимально приближена к остальным зубам. Оттенок окружающих зубов будет записан, чтобы убедиться, что цвет выглядит естественным и соответствует этим зубам.
Будет ли корона ощущаться иначе?Поскольку форма коронки будет немного отличаться от формы вашего зуба до установки коронки, вы можете сразу это заметить.Через несколько дней он должен чувствовать себя хорошо, и вы этого не заметите. Коронке может потребоваться некоторая корректировка, если ваш прикус неудобен, и в этом случае вам следует попросить своего стоматолога проверить и отрегулировать ее.
Сколько это будет стоить?Стоимость зависит от типа коронки и используемого материала. Всегда получайте письменную оценку и план лечения перед началом любого стоматологического лечения.
Как ухаживать за короной?Важно, чтобы коронка была такой же чистой, как и ваши естественные зубы.Сама коронка не может разрушаться, но разрушение может начаться там, где край коронки соединяется с зубом. В последнюю очередь чистите зубы на ночь и, по крайней мере, один раз в течение дня зубной пастой с фтором, а также очищайте межзубные промежутки межзубными щетками или зубной нитью.
Как долго прослужит корона?Срок службы вашей короны зависит от того, насколько хорошо вы за ней ухаживаете. Коронки, за которыми правильно ухаживают, должны служить долгие годы. Ваш стоматолог сможет сказать вам, как долго может прослужить ваша коронка.
Люди, просматривавшие эту страницу, также посещали…
Нужен совет?
Если вам нужен бесплатный и беспристрастный совет по поводу здоровья полости рта, свяжитесь с нашей службой стоматологической помощи по электронной почте или по телефону 01788 539780 (звонок по местному тарифу в Великобритании).
Наша служба стоматологической помощи является полностью конфиденциальной и помогла почти 400 000 человек с момента открытия более 20 лет назад. Свяжитесь с нашими экспертами по телефону, электронной почте или через Интернет с понедельника по пятницу с 09:00 до 17:00.
Опубликовано:
Обновлено:
Автор: Джордж Бушелл
.