У фасоли корневая система стержневая
Фасоль обыкновенная
Фасоль обыкновенная( Phaseolus Vlgaris)
Это однолетнее, кустовое, сильно вьющееся, его плети достигают до 70 сантиметров в длину, растение, семейства бобовых. Фасоль является древнейшим культурным растением. Ее родиной принято считать Южную Америку, где ее с древних времен выращивали индейцы, и она являлась одним из важнейших культур земледелия. Но есть сведения о том, что о фасоли есть упоминания и в летописях Древнего Египта и Древнего Рима. Древние римляне не только употребляли фасоль в пищу, но и использовали ее для приготовления пищи. В Европе, как пищевое и лекарственное растение фасоль появилась в середине ХVI века, об этом есть указания в травниках тех времен. Откуда попала фасоль в Россию не известно. Есть предположения, что впервые привезли ее из Франции, так как очень долгое время фасоль называли «французскими бобами». В начале в России фасоль разводилась, как декоративное растение, и лишь в ХVII веке фасоль получила широкое распространение, как пищевая культура.
Фасоль является ценной продовольственной культурой. По своему составу белки фасоли не на много уступают белкам мяса и очень хорошо усваиваются организмом человека. На сегодняшний день фасоль занимает второе место по популярности среди бобовых продовольственных культур. Для многих народов южных стран фасоль является главнейшей продовольственной культурой.
Корневая система: корень фасоли стержневой, сильно разветвленный. Большая часть корневой системы расположена под верхними слоями почвы, но существуют корни, которые уходит в землю на глубину до 75 сантиметров. Фасоль имеет одно очень замечательное свойство, она подобно гороху и другим бобовым способна извлекать азот из воздуха, тем самым обогащая почву. После уборки урожая корни растения оставляют в почве, как органическое удобрение.
Стебель:
У некоторых сортов фасоли стебель травянистый вьющейся, у других – прямой сильно разветвленный покрытый редкими жесткими волосками.
Листья парноперистые на длинных черешках.
Цветки фасоли бываю белого, розового и лилового цвета. Собраны в короткие пазушные кисти по 2-6 штук на длинных цветоножках.
Лекарственное сырье:
В лечебных целях используют хорошо вызревшие и пожелтевшие створки стручков фасоли. В некоторых случаях в народной медицине, как лекарственное средство используются цветки и околоплодники. Сырье (створки стручков) обладает антидиабетическим действием. В зрелых плодах фасоли содержаться аминокислоты, аргинин, тригонеллин, пиразин, лейцин, лизин, триптофан, холин, аспарагин и гемицеллюлоза, органические и минеральные кислоты, соли фосфора и калия, аскорбиновую кислоту и витамины группы. Аргинин, содержащийся в бобах фасоли оказывает инсулиноподобное действие на обмен веществ в организме.
Сбор и заготовки: Для лечебных целей используют сухие створки стручков, после их полного созревания. Собирают их в августе в начале сентября. Есть разновидности фасоли, у которой в пищу употребляют в пищу молодые незрелые, так называемые флажолеты, стручки.
Есть разновидности фасоли, у которой в пищу употребляют в пищу молодые незрелые, так называемые флажолеты, стручки.
Применение:
Фасоль содержит в себе целый список полезных веществ и минералов, полезных для организма человека. Но к сожалению в научной медицине она не нашла своего применения.
В народной же медицине разных стран, ее целебные качества используются с древних времен. Водные настои и отвары из стручков используется для лечения различных заболеваний почек и мочевого пузыря, гипертонии, сердечно сосудистой слабости, хроническом ревматизме, подагре.
Отвар из сушеных цветков фасоли пьют при камнях в почках. В народе распространено употреблении муки из фасоли для присыпки различных видов экзем и кожной сыпи. Водный раствор из створок применяют при отеках вызванных болезнью почек. При ревматизме рекомендуют пить отвар из околоплодников. Фасоль является немаловажным продуктом питания в рационе больных сахарным диабетом. Употребление в пищу фасоли, способствует снижению содержания сахара в крови.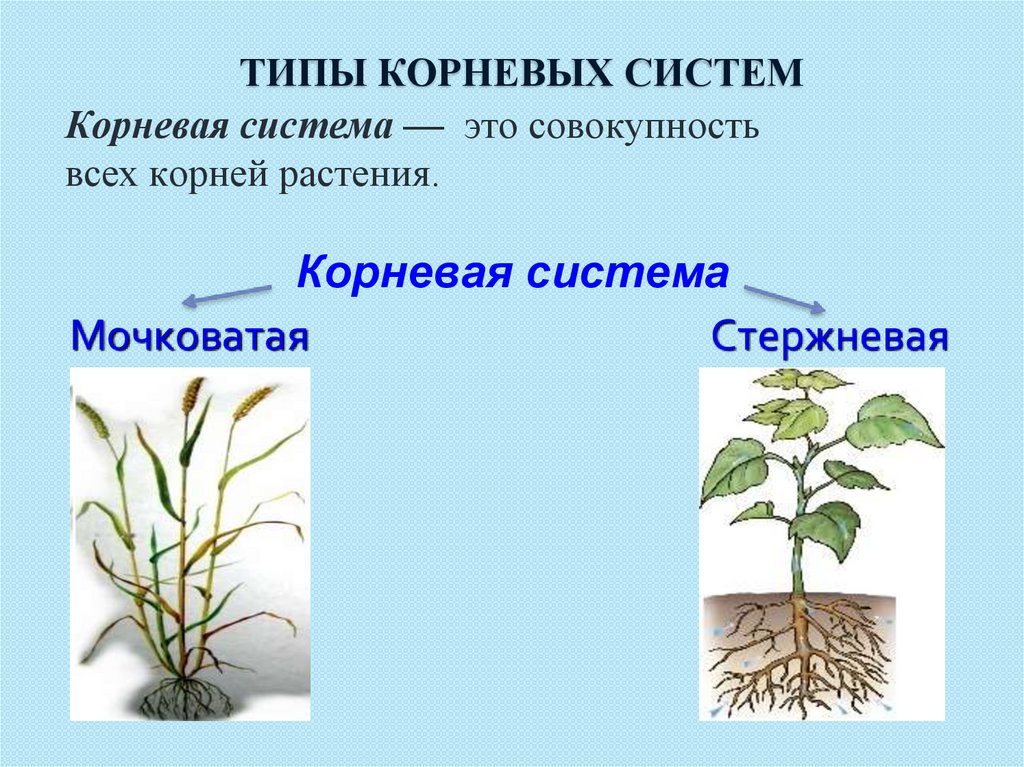
Овощная фасоль усиливает секрецию желудочного сока. Пюре из фасоли включены в рационы как диетические блюда для больных гастритами и заболевания печени, но употребление его должно быть несколько ограниченным, так как некоторые вещества могут вызывать вздутие кишечника.
Главная причина неудач в выращивании растения фасоли заключается в слабом знании её биологических особенностей. Поэтому любому дачнику, планирующему освоить агротехнику этой полезной и вкусной культуры, я советую начинать знакомство с краткого биопортрета.По типу роста растение фасоли бывает вьющееся и кустовое.
Стебель у него травянистый, тёмно-зелёный, розоватый или фиолетовый. Длина стебля у кустовых сортов составляет от 25 до 60 сантиметров, а у вьющихся может достигать до 5 метров (в среднем — 2-3 метра).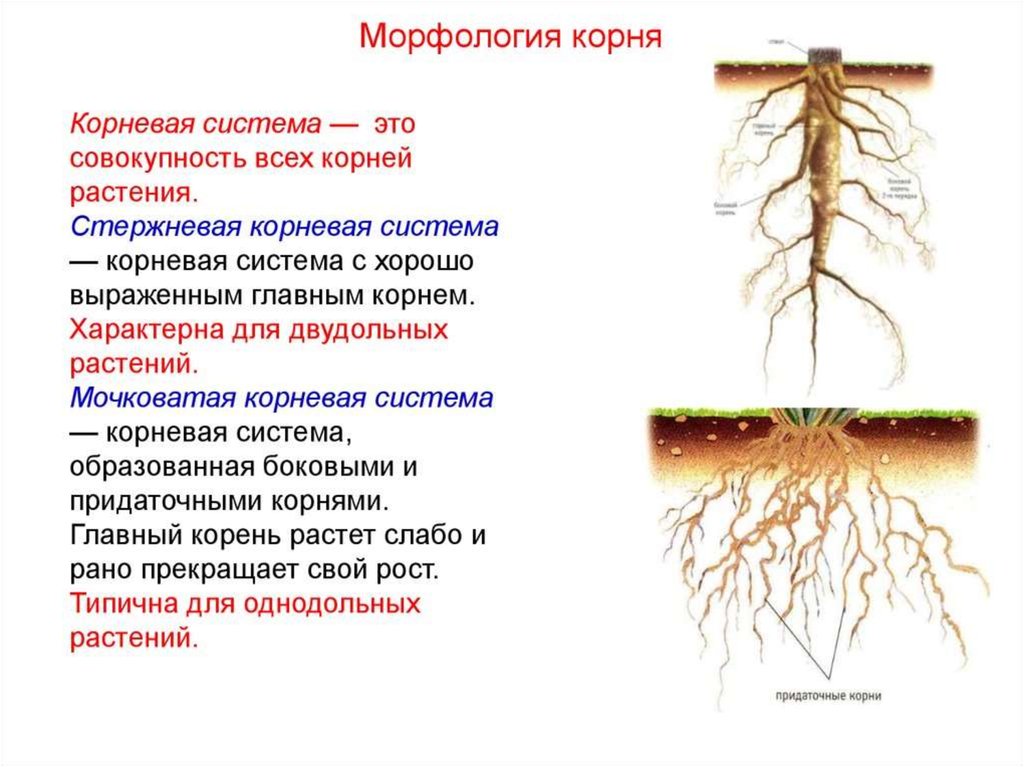 Это позволяет использовать растение для вертикального озеленения.
Это позволяет использовать растение для вертикального озеленения.
Кустовые сорта фасоли различаются по общему размеру куста: он может быть как довольно компактным, так и раскидистым.
Корневая система у фасоли имеет стрежневое строение и проникает у вьющихся сортов на метр в толщу почвы. При этом она занимает площадь в радиусе от 55 до 60 сантиметров.
У кустовой фасоли корни развиты скромнее и залегают на глубине порядка 20-30 сантиметров. На той же глубине расположен и стержневой корень. Боковые корешки плотные и не уступают ему по протяжённости.
На корнях фасоли развиваются особая разновидность клубеньковых бактерии, которые способны при участии растения извлекать из воздуха азот и насыщать им окружающую почву. При хороших условиях на корневой системе одного фасолевого куста формируется от 500 до 700 клубеньков! В итоге после уборке ботвы овоща остаётся обогащённый азотом грунт. За сезон в нём аккумулируется до 44 грамм макроэлемента на 10 квадратных метра.
Листья у фасоли сложные, тройчатообразные с большими заострёнными или овальными листочками. Первая пара листочков имеет простую форму, а все последующие — непарноперистые. Прилистники мелкие яйцеоподобные или ланцетные. Листовая пластинка окрашена в жёлто-зелёный или насыщенно-зелёный с фиолетовым отливом цвет.
Фасолевые цветки мотылькового вида, некрупные, состоят из 5 лепестков и организованы в соцветие-кисть. Цветение кустовых сортов длится от 20 до 25 дней, вьющихся от 1 до 3 месяцев. Цветоносы укороченные, верхушечные либо пазушные, чаще с парным размещением цветков.
У многоцветковых сортов фасоли цветоносы вырастают до большой длины и цветки формируются над листьями. Их длина обычно составляет около 45 сантиметров, а количество цветков составляет от 40 и более.
Окраска цветков может быть розовая, белая, ярко-красная или двухцветная. Зачастую у сортов со светлыми цветками семена белые, с розовыми — коричневые или красновато-коричневые, а с фиолетовыми – т ёмные, почти чёрные.
Плод у растения фасоли представляет собой боб (лопатку) длиной от 8 до 25 сантиметров и содержит до 12 семян самой разной формы и цвета. По форме они бывают прямые, саблевидные, изогнутые, серповидные, мечевидные либо циллиндрические.
Масса 1000 небольших семян фасоли не превышает 300 грамм, а крупных — не более 450 грамм.
Согласно археологическим находкам, в древности растение фасоль возделывали на территории тропической Америки. Семена растения были обнаружены в ископаемых останках ранних мексиканских и перуанских народностей.
Фасоль вместе с тыквой и кукурузой составляли триаду важнейших культурных растений древней Мексики. Причём фасоль и кукуруза выращивались в совместных посадках: кукуруза играла роль опоры для вьющихся разновидностей фасоли. Словом, наши дальние предки знали толк в интенсивном земледелии.
Для растения фасоли характерна большая изменчивость, то есть существует немало видов и подвидов культуры.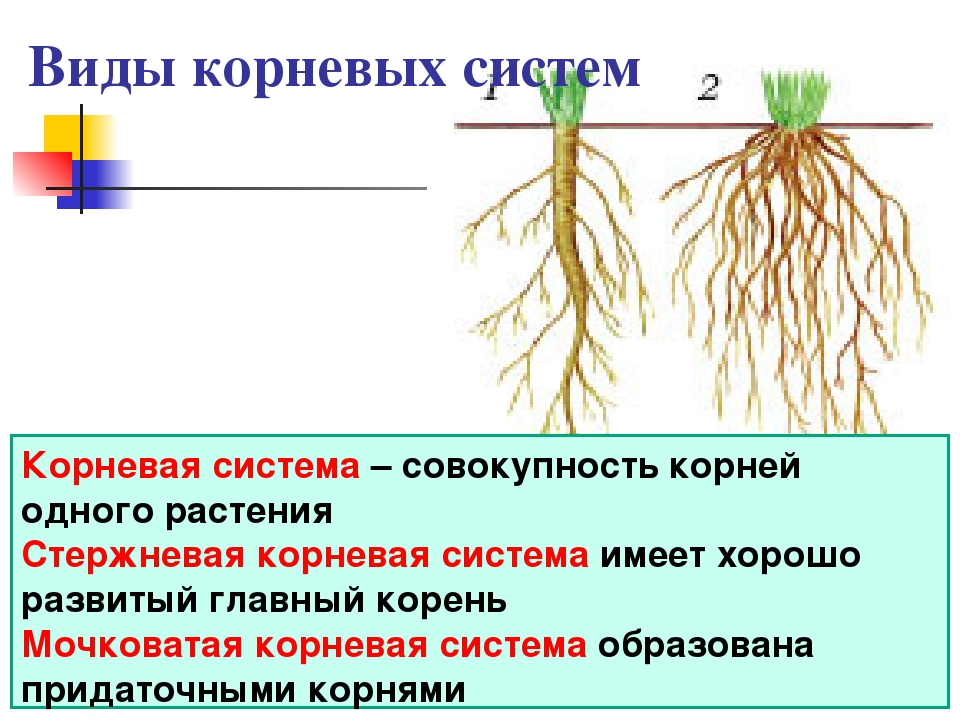
На Родине она встречается как в форме однолетника, так и в виде двулетника и даже многолетника. Там она представлена лианоподобными, вьющимися и стелющимися сортами с огромным количеством боковых побегов, переплетающихся между собой. Кустовые разновидности фасоли появились в результате спонтанных мутаций оригинальных видов и были введены в культуру стараниями человека.
Из тропической Америки фасоль была завезена испанскими путешественниками в Европу.
Первое время её возделывали исключительно в качестве декоративного растения. Однако в XVII веке, когда стало известно о питательной ценности фасолевых бобов, она быстро стала популярной во всех европейских странах.
В России фасоль также культивируют уже довольно давно, но у нас больше распространена зерновая фасоль, у которой в еду используются вызревшие семена. В нашу страну фасоль попала из Турции и Грузии, поэтому вначале ее называли турецкими бобами. Спаржевая фасоль пока не заслужила широкого признания среди российских дачников.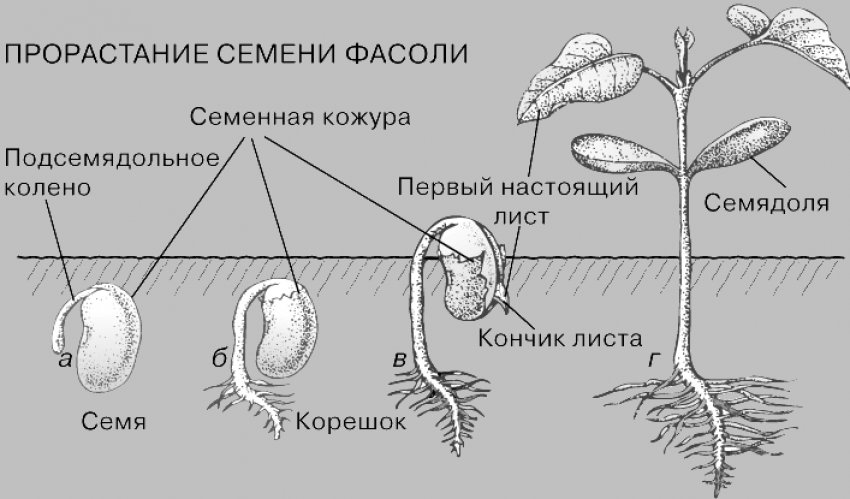 Более подробно о разных видах фасоли читайте здесь.
Более подробно о разных видах фасоли читайте здесь.
Какую фасоль мы сажаем у себя на участке, я рассказала и показала в видео.
Хорошо известная нам фасоль является типичным растением из семейства бобовых. Этот род растений был завезен к нам из стран Южной Америки после открытия континента Колумбом. Из статьи вы узнаете, как образуются семена фасоли, об их строении, составе и калорийности.
Как образуются семена фасоли?
Семена этого бобового растения появляются в результате оплодотворения цветка опылением, образованием завязи из нижней части пестика. У фасоли это происходит самоопылением, изредка при помощи насекомых происходит перекрёстное опыление. Каждый цветок цветёт 2–3 суток, кисть — 10–14 дней. Цветение одного растения длится до 25 суток.
Описание семян и их значение
Семена фасоли типичны для покрытосеменного двудольного растения. Они имеют форму бобов с выпуклой и вогнутой сторонами. Вогнутая часть имеет заметный рубчик — место крепления семяножки, связующей с околоплодником. Рядом с ним наблюдается мелкое отверстие — семявход (микропиле), через которое внутрь проникает влага и воздух. Цвет зависит от сортовых или видовых особенностей и может быть белым, коричневым, лиловых и других тонов.
Они могут быть не только однотонными, но и в крапинку, мозаичными, пятнистыми. Семечко фасоли, по сути, является зачатком молодого растения, защищённого семенной кожурой, имеющего запас питания на первое время развития. Питательные вещества находятся в эндосперме.
Питательные вещества находятся в эндосперме.Зародыш
При удалении кожуры с боба можно увидеть зародыш. Он находится рядом с микропиле, и является основой будущего молодого растения. По схеме, в его строении различают, стебелёк, почечку, семядоли и зародышевый корешок — те же части, что и у взрослого растения.
Зародышевый корешок
Корешок зародыша — это зачаток будущей корневой системы. Он впоследствии формируется в главный (стержневой) корень растения. Находится ближе всех к семявходу и первым выбирается наружу. Корень проростка закрепляется в грунте, начинает добычу воды и питания.
Семядоли
Фасоль относится к двудольным растениям. Это значит, что у её зародыша имеются две супротивные боковые семядоли. У этого рода растений они толстые и мясистые. Это первая эмбриональная пара листьев будущего растения. Они содержат немало питательных веществ для ростка, взятые из эндосперма. Также листья защищают почечку, что немаловажно при пробивании ростка сквозь корку грунта. Когда росток наберёт сил, семядоли засохнут, предварительно отдав ему накопленные питательные вещества.
Когда росток наберёт сил, семядоли засохнут, предварительно отдав ему накопленные питательные вещества.
Что необходимо для прорастания зародыша?
Всё, что нужно для прорастания — это тепло, влага, воздух и свет. Фасолине при проращивании нужно много влаги (100–120%). Минимальная температура прорастания семян фасоли — это +8…+10°С. Чем выше температура, тем быстрее появляются всходы. Оптимальной считается температура +20. +22°С. При лучшем температурном режиме всходы появляются на 6–8 сутки. Они могут погибнуть при похолодании до –0,5…+1°С.
Семечко не должно находиться полностью погружённым в воду или в слишком плотном грунте, иначе не будет поступления воздуха. При нехватке света росток будет чахлым и бледным, а со временем погибнет. Не взойдут семена, повреждённые вредителями и болезнями, с истёкшим сроком годности. Срок годности посевных семян этой культуры 5–7 лет. Лучшая вода для проращивания семян — талая.Химический состав и калорийность
Фасоль является ценным источником питания.
В 100 г находится около 298 ккал, а также:
- углеводов 47 г;
- белков 21 г;
- жиров 2 г;
- пищевых волокон 12,4 г;
- воды 14 г;
- золы 3,6 г.
Витамины:
- РР — 6,4 мг;
- ниацин — 2,1 мг;
- В5 — 1,2 мг;
- В6 — 0,9 мг;
- Е — 0,6 мг;
- В1 — 0,5 мг;
- В2 — 0,18 мг;
- В9 — 0,09 мг.
Минералы:
- калий 1,100 г;
- фосфор 480 мг;
- сера 159 мг;
- кальций 150 мг;
- магний 103 мг;
- кремний 92 мг;
- натрий 40 мг;
- хлор 58 мг;
- железо 5,9 мг;
- марганец 1,34 мг;
- цинк 3,21 мг;
- алюминий 0,64 мг;
- медь 0,58 мг;
- бор 0,49 мг;
- ванадий 0,19 мг;
- никель 0,1732 мг;
- титан 0,15 мг;
- фтор 44 мкг;
- молибден 39,4 мкг;
- селен 24,9 мкг;
- кобальт 18,7 мкг;
- йод 12,1 мкг;
- хром 10 мкг.

Поскольку в фасоли немало витаминов группы В, особенно фолатов, это важно для диетического питания и правильного формирования плода у беременных. Эти бобы содержат 12 незаменимых аминокислот и 8 заменимых. Вареные зёрна в зависимости от разновидности содержат уже 118–147 ккал. При рационе с регулярным потреблением этого бобового снижается уровень холестерина, нормализуется уровень сахара, давление, успокаиваются нервы, укрепляются кости и увеличивается энергичность. Семечко фасоли содержит зачаток будущего растения и прорастёт при подходящих для этого условиях. Оно имеет большую пищевую ценность.
особенности и описание, строение корневой системы, семени
Хорошо известная нам фасоль является типичным растением из семейства бобовых. Этот род растений был завезен к нам из стран Южной Америки после открытия континента Колумбом. Из статьи вы узнаете, как образуются семена фасоли, об их строении, составе и калорийности.
ПоказатьСкрытьКак образуются семена фасоли?
Семена этого бобового растения появляются в результате оплодотворения цветка опылением, образованием завязи из нижней части пестика. У фасоли это происходит самоопылением, изредка при помощи насекомых происходит перекрёстное опыление. Каждый цветок цветёт 2–3 суток, кисть — 10–14 дней. Цветение одного растения длится до 25 суток. У овощной культуры при повторных сборах молодых лопаток время цветения вьющихся разновидностей может составить от 30 до 100 суток. Пыльца у фасоли обыкновенной вызревает ещё в нераскрывшемся цветке, и самоопыляется культура за несколько часов до раскрытия лепестков. В каждом двустворчатом стручке фасоли обыкновенной (самый распространённый вид) образовывается от 2 до 8 семян.
У фасоли это происходит самоопылением, изредка при помощи насекомых происходит перекрёстное опыление. Каждый цветок цветёт 2–3 суток, кисть — 10–14 дней. Цветение одного растения длится до 25 суток. У овощной культуры при повторных сборах молодых лопаток время цветения вьющихся разновидностей может составить от 30 до 100 суток. Пыльца у фасоли обыкновенной вызревает ещё в нераскрывшемся цветке, и самоопыляется культура за несколько часов до раскрытия лепестков. В каждом двустворчатом стручке фасоли обыкновенной (самый распространённый вид) образовывается от 2 до 8 семян.
Знаете ли вы? Перекрёстно опыляется большинство цветков многоцветкового вида. Этот вид ещё называют турецкими бобами. Он красиво цветёт и нередко используется как декоративное растение. В пищу пригодны только молодые бобы.
Описание семян и их значение
Семена фасоли типичны для покрытосеменного двудольного растения. Они имеют форму бобов с выпуклой и вогнутой сторонами.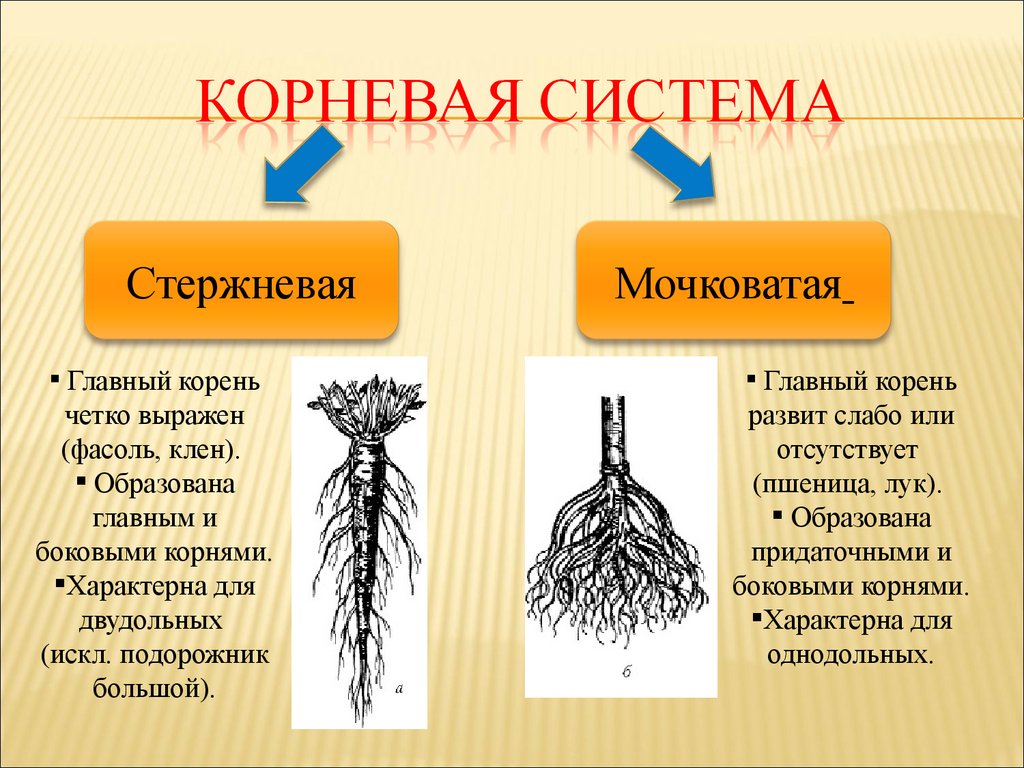 Вогнутая часть имеет заметный рубчик — место крепления семяножки, связующей с околоплодником. Рядом с ним наблюдается мелкое отверстие — семявход (микропиле), через которое внутрь проникает влага и воздух. Цвет зависит от сортовых или видовых особенностей и может быть белым, коричневым, лиловых и других тонов. Они могут быть не только однотонными, но и в крапинку, мозаичными, пятнистыми. Семечко фасоли, по сути, является зачатком молодого растения, защищённого семенной кожурой, имеющего запас питания на первое время развития. Питательные вещества находятся в эндосперме.
Вогнутая часть имеет заметный рубчик — место крепления семяножки, связующей с околоплодником. Рядом с ним наблюдается мелкое отверстие — семявход (микропиле), через которое внутрь проникает влага и воздух. Цвет зависит от сортовых или видовых особенностей и может быть белым, коричневым, лиловых и других тонов. Они могут быть не только однотонными, но и в крапинку, мозаичными, пятнистыми. Семечко фасоли, по сути, является зачатком молодого растения, защищённого семенной кожурой, имеющего запас питания на первое время развития. Питательные вещества находятся в эндосперме.
Знаете ли вы? Самые большие семена формирует веерная пальма — это 25-килограммовый орех. Он прорастает целых три года.
Зародыш
При удалении кожуры с боба можно увидеть зародыш. Он находится рядом с микропиле, и является основой будущего молодого растения. По схеме, в его строении различают, стебелёк, почечку, семядоли и зародышевый корешок — те же части, что и у взрослого растения.
Зародышевый корешок
Корешок зародыша — это зачаток будущей корневой системы. Он впоследствии формируется в главный (стержневой) корень растения. Находится ближе всех к семявходу и первым выбирается наружу. Корень проростка закрепляется в грунте, начинает добычу воды и питания.
Семядоли
Фасоль относится к двудольным растениям. Это значит, что у её зародыша имеются две супротивные боковые семядоли. У этого рода растений они толстые и мясистые. Это первая эмбриональная пара листьев будущего растения. Они содержат немало питательных веществ для ростка, взятые из эндосперма. Также листья защищают почечку, что немаловажно при пробивании ростка сквозь корку грунта. Когда росток наберёт сил, семядоли засохнут, предварительно отдав ему накопленные питательные вещества.
Важно! Фасоль не стоит употреблять пожилым людям, а также при проблемах с ЖКТ (повышенная кислотность, язвенные болезни, гастрит), при подагре, холецистите и болезнях поджелудочной железы.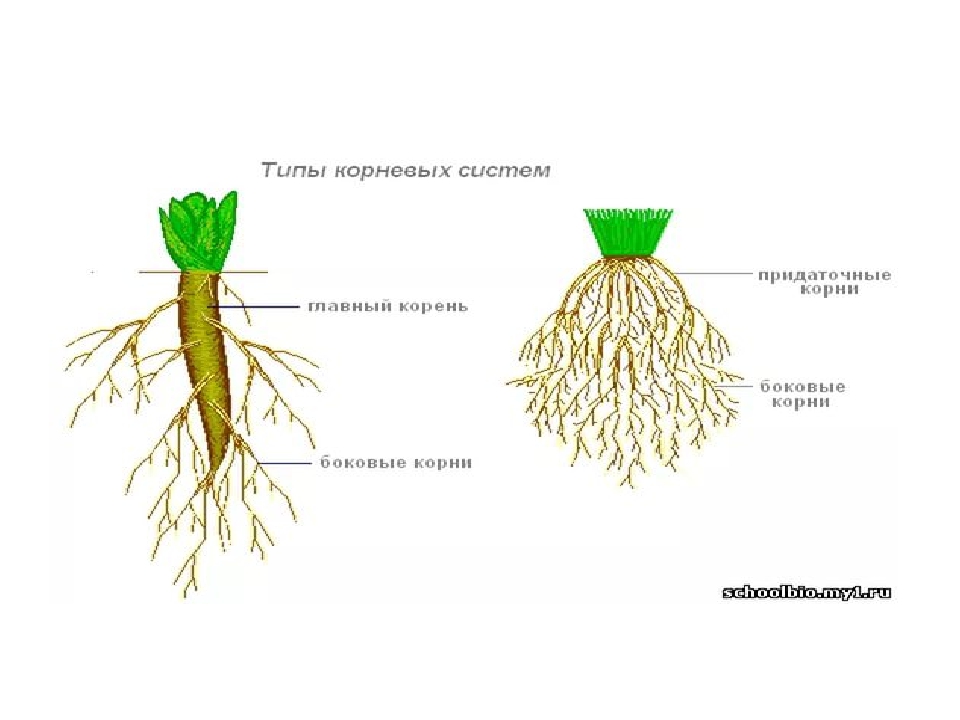
Что необходимо для прорастания зародыша?
Всё, что нужно для прорастания — это тепло, влага, воздух и свет. Фасолине при проращивании нужно много влаги (100–120%). Минимальная температура прорастания семян фасоли — это +8…+10°С. Чем выше температура, тем быстрее появляются всходы. Оптимальной считается температура +20…+22°С. При лучшем температурном режиме всходы появляются на 6–8 сутки. Они могут погибнуть при похолодании до –0,5…+1°С. Семечко не должно находиться полностью погружённым в воду или в слишком плотном грунте, иначе не будет поступления воздуха. При нехватке света росток будет чахлым и бледным, а со временем погибнет. Не взойдут семена, повреждённые вредителями и болезнями, с истёкшим сроком годности. Срок годности посевных семян этой культуры 5–7 лет. Лучшая вода для проращивания семян — талая.
Химический состав и калорийность
Фасоль является ценным источником питания.
В 100 г находится около 298 ккал, а также:
- углеводов 47 г;
- белков 21 г;
- жиров 2 г;
- пищевых волокон 12,4 г;
- воды 14 г;
- золы 3,6 г.
Витамины:
- РР — 6,4 мг;
- ниацин — 2,1 мг;
- В5 — 1,2 мг;
- В6 — 0,9 мг;
- Е — 0,6 мг;
- В1 — 0,5 мг;
- В2 — 0,18 мг;
- В9 — 0,09 мг.
Минералы:
- калий 1,100 г;
- фосфор 480 мг;
- сера 159 мг;
- кальций 150 мг;
- магний 103 мг;
- кремний 92 мг;
- натрий 40 мг;
- хлор 58 мг;
- железо 5,9 мг;
- марганец 1,34 мг;
- цинк 3,21 мг;
- алюминий 0,64 мг;
- медь 0,58 мг;
- бор 0,49 мг;
- ванадий 0,19 мг;
- никель 0,1732 мг;
- титан 0,15 мг;
- фтор 44 мкг;
- молибден 39,4 мкг;
- селен 24,9 мкг;
- кобальт 18,7 мкг;
- йод 12,1 мкг;
- хром 10 мкг.
Поскольку в фасоли немало витаминов группы В, особенно фолатов, это важно для диетического питания и правильного формирования плода у беременных. Эти бобы содержат 12 незаменимых аминокислот и 8 заменимых. Вареные зёрна в зависимости от разновидности содержат уже 118–147 ккал. При рационе с регулярным потреблением этого бобового снижается уровень холестерина, нормализуется уровень сахара, давление, успокаиваются нервы, укрепляются кости и увеличивается энергичность. Семечко фасоли содержит зачаток будущего растения и прорастёт при подходящих для этого условиях. Оно имеет большую пищевую ценность.
Вареные зёрна в зависимости от разновидности содержат уже 118–147 ккал. При рационе с регулярным потреблением этого бобового снижается уровень холестерина, нормализуется уровень сахара, давление, успокаиваются нервы, укрепляются кости и увеличивается энергичность. Семечко фасоли содержит зачаток будущего растения и прорастёт при подходящих для этого условиях. Оно имеет большую пищевую ценность.
Урок13.1. Практическая работа 9. Типы корневых систем. Строение стержневой и мочковатой корневых систем
Методическое пособие разработки уроков биологии 6класс
Тип урока — комбинированный
Методы: частично-поисковый, проблемного изложения, репродуктивный, объяснительно-иллюстративный.
Цель:
— осознание учащимися значимости всех обсуждаемых вопросов, умение строить свои отношения с природой и обществом на основе уважения к жизни, ко всему живому как уникальной и бесценной части биосферы;
Задачи:
Образовательные: показать множественность факторов, действующих на организмы в природе, относительность понятия «вредные и полезные факторы», многообразие жизни на планете Земля и варианты адаптаций живых существ ко всему спектру условий среды обитания.
Развивающие: развивать коммуникативные навыки, умения самостоятельно добывать знания и стимулировать свою познавательную активность; умения анализировать информацию, выделять главное в изучаемом материале.
Воспитательные:
Формирование экологической культуры на основе признания ценности жизни во всех её проявлениях и необходимости ответственного, бережного отношения к окружающей среде.
Формирование понимания ценности здорового и безопасного образа жизни
УУД
Личностные:
воспитание российской гражданской идентичности: патриотизма, любви и уважения к Отечеству, чувства гордости за свою Родину;
Формирование ответственного отношения к учению;
3) Формирование целостного мировоззрения, соответствующего современному уровню развития науки и общественной практики.
Познавательные: умение работать с различными источниками информации, преобразовывать её из одной формы в другую, сравнивать и анализировать информацию, делать выводы, готовить сообщения и презентации.
Регулятивные: умение организовать самостоятельно выполнение заданий, оценивать правильность выполнения работы, рефлексию своей деятельности.
Коммуникативные: Формирование коммуникативной компетентности в общении и сотрудничестве со сверстниками, старшими и младшими в процессе образовательной, общественно полезной, учебно-исследовательской, творческой и других видов деятельности.
Планируемые результаты
Предметные: знать — понятия «среда обитания», «экология», «экологические факторы» их влияние на живые организмы, «связи живого и неживого»;. Уметь — определять понятие «биотические факторы»; характеризовать биотические факторы, приводить примеры.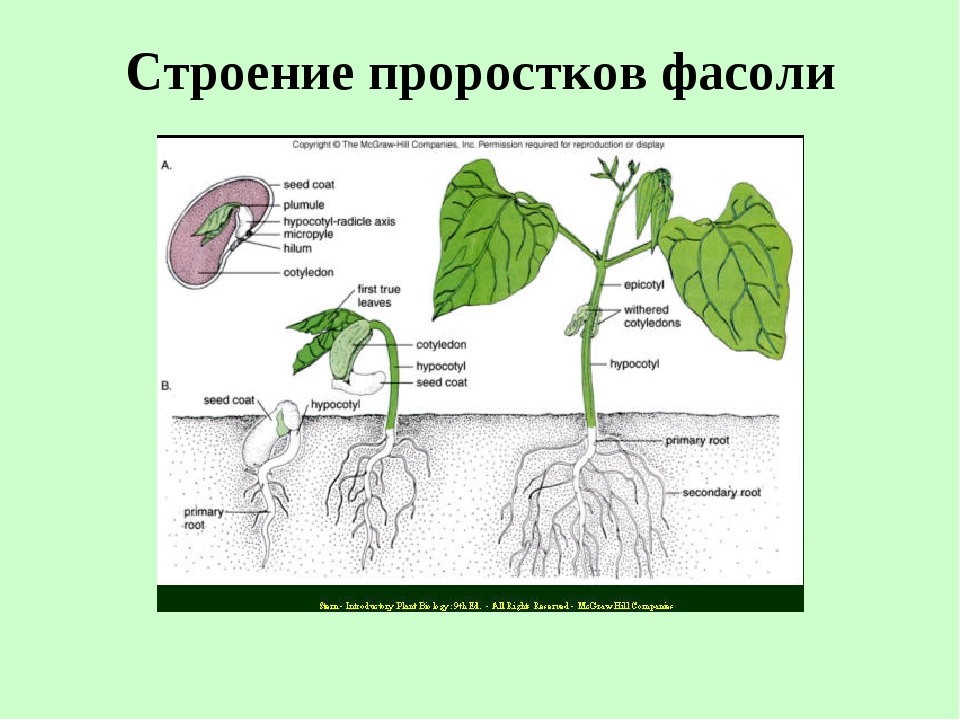
Личностные: высказывать суждения, осуществлять поиск и отбор информации; анализировать связи, сопоставлять, находить ответ на проблемный вопрос
Метапредметные:.
Умение самостоятельно планировать пути достижения целей, в том числе альтернативные, осознанно выбирать наиболее эффективные способы решения учебных и познавательных задач.
Формирование навыка смыслового чтения.
Форма организации учебной деятельности – индивидуальная, групповая
Методы обучения: наглядно-иллюстративный, объяснительно-иллюстративный, частично-поисковый, самостоятельная работа с дополнительной литературой и учебником, с ЦОР.
Приемы: анализ, синтез, умозаключение, перевод информации с одного вида в другой, обобщение.
Цель: познакомить со строением и развитием стержневых и мочковатых корневых систем.
Оборудование: проростки семян фасоли (гороха, огурца и т. д.), зерновок пшеницы (ржи, кукурузы и т. д.) в возрасте 5—7 и 10—12 дней; гербарии растений со стержневой и мочковатой корневыми системами.
Ход работы
Рассмотрите проросшие семена фасоли 5—7-дневного возраста. Какой корень вы видите?
Рассмотрите проросшие семена фасоли 10—12-дневного возраста. Появились ли какие-нибудь изменения в корневой системе?
Рассмотрите корневую систему взрослого растения фасоли. Найдите главный и боковые корни. Как называется такой тип корневой системы?
Зарисуйте корневую систему проростка фасоли в возрасте 5—7 и 10—12 дней, а также корневую систему взрослого растения фасоли. Подпишите тип корневой системы и все виды корней.
Рассмотрите проросшие зерновки пшеницы 5—7-дневного возраста. Сколько корней появляется на первой стадии развития проростка пшеницы?
Сколько корней появляется на первой стадии развития проростка пшеницы?
Рассмотрите проросшие зерновки пшеницы 10—12-дневного возраста. Как происходит формирование корневой системы пшеницы? Какие типы корней вы видите? В чем отличие развития корневых систем пшеницы и фасоли?
Рассмотрите корневую систему взрослого растения пшеницы. Можете ли вы отличить главный корень? Найдите придаточные и боковые корни.
Зарисуйте корневую систему проростка пшеницы в возрасте 5—7 и 10—12 дней, а также корневую систему взрослого растения пшеницы. Подпишите тип корневой системы и все виды корней.
Сравните корневые системы фасоли и пшеницы.
Сделайте вывод о сходствах и различиях корневых систем фасоли и пшеницы.
Рассказ учителя с элементами беседы
Размер и форма корневой системы растений зависят и от условий их произрастания. Например, корневая система сосны, растущей на песчаной почве, будет отличаться от корневой системы сосны, растущей на болоте.
Например, корневая система сосны, растущей на песчаной почве, будет отличаться от корневой системы сосны, растущей на болоте.
Подумайте, как будет выглядеть корневая система дерева в том и другом случае. (Ответы учащихся.)
У сосны, растущей на песчаной почве, корневая система будет стержневой, как и положено двудольному растению. Кроме того, корни его будут проникать глубоко в почву в поисках влаги. Сосна, растущая на болоте, будет иметь корневую систему, больше напоминающую мочковатую. А ее корни будут занимать большую площадь, но не проникнут глубоко в землю.
Как вы думаете, почему? (Это необходимо растению, чтобы удержаться на неплотной болотистой почве.)
Стержневую корневую систему растения можно изменить, превратив ее в мочковатую. Корень растет верхушкой.
Но что будет с корнем, если мы удалим верхушку? (Ответы учащихся.)
Если удалить верхушку главного корня, то его рост прекратится, зато боковые корни начнут отрастать быстрее. Это свойство корней используют в растениеводстве. При пересадке рассады в открытый грунт прищипывают кончик главного корня. Этот метод называется прищипывание, или пикировка.
Это свойство корней используют в растениеводстве. При пересадке рассады в открытый грунт прищипывают кончик главного корня. Этот метод называется прищипывание, или пикировка.
Как вы думаете, для чего прищипывают корни сельскохозяйственных растений? (Ответы учащихся.)
В результате прищипывания развивается сильно разветвленная корневая система, которая располагается в верхних, самых плодородных слоях почвы. Соответственно, проросток будет развиваться быстрее, а в результате растение даст больший урожай.
Какие функции выполняют корни растений? (Ответы учащихся.)
Кроме основных — водно-солевого питания и укрепления в почве, — у корней есть и другое назначение. Например, у некоторых растений в корнях могут откладываться запасные питательные вещества. Эти корни сильно разрастаются в толщину и приобретают иногда очень диковинную форму.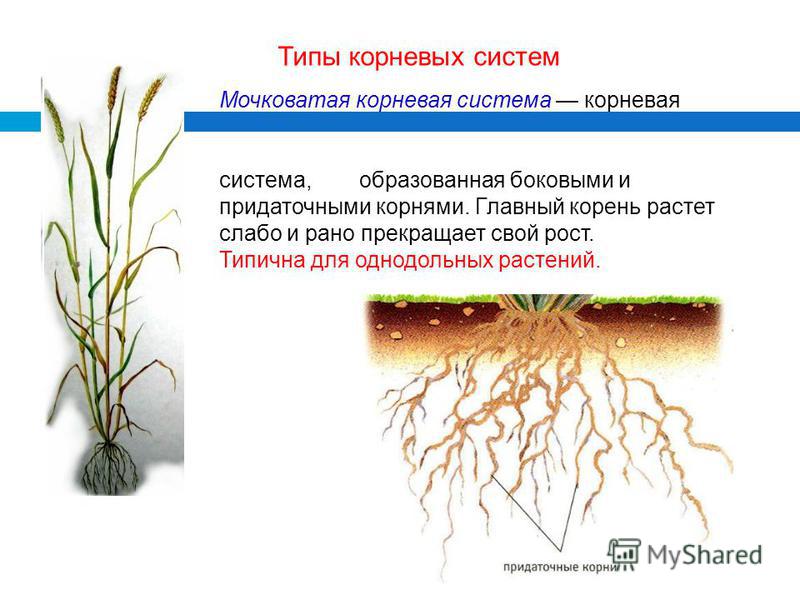
У каких растений питательные вещества запасаются в корнях? (Ответы учащихся.)
Такие видоизменения корней называют корнеплодами, они образуются из главного корня и нижних участков стебля. В корнеплодах может запасаться сахар, тогда они имеют сладкий вкус.
Корнеплоды каких растений сладкие? (Моркови, сахарной свеклы.)
Растения, образующие корнеплоды, чаще всего являются двулетниками.
Вспомните, как развиваются двулетники в первый и во второй год. (В первый год они не цветут, а накапливают питательные вещества. Во второй — используют запасенные питательные вещества, цветут и плодоносят.)
Некоторые растения образуют не один корнеплод, а множество корневых клубней, возникающих на боковых или придаточных корнях. Корневые клубни образуют георгин, чистяк, батат. (Батат — растение, возделываемое в тропических и субтропических странах и заменяющее местным жителям картофель.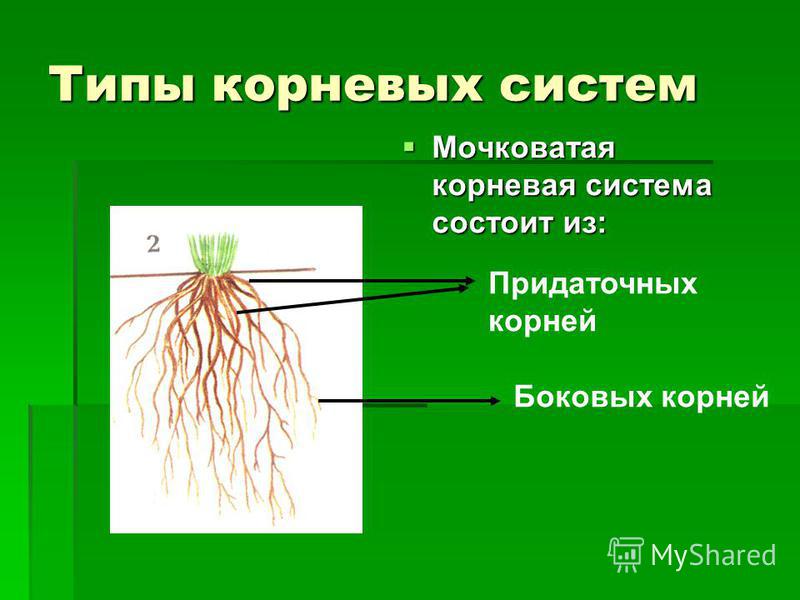 )
)
У многих растений семейства бобовых на боковых корнях образуются особые бактериальные клубеньки.
В молодых корнях этих растений поселяются особые бактерии, которые способствуют усвоению азота из воздуха, тем самым давая возможность растениям благополучно обитать на бедных азотом почвах. Кроме того, азот накапливается в самих корнях этих растений, обогащая почву.
У некоторых луковичных растений (гусиный лук, крокусы, гладиолусы) корни выполняют еще одну очень специфическую функцию. Отдельные корешки устроены таким образом, что, сокращаясь, подобно растянутой резинке, способны втягивать луковицу и увлекать ее за собой глубже под землю.
У некоторых тропических растений, как, например, у тропических орхидей, имеются особые придаточные корни. Они приспособились получать влагу из влажного воздуха, поэтому их называют воздушными корнями.
(В кабинетах биологии часто встречается растение монстера, на примере которого можно продемонстрировать детям воздушные корни. )
)
Как мы уже говорили раньше, все органы растения нуждаются в кислороде для дыхания. Но в заболоченных почвах воздуха крайне мало, так как все свободное пространство между частичками почвы заполнено водой. Такие условия характерны для мангровых зарослей, поэтому корни многих из обитающих здесь растений поднимаются над поверхностью почвы и поглощают кислород из воздуха, т. е. дышат. Эти корни так и называются — дыхательные.
Растения мангровых зарослей используют придаточные корни не только для питания или дыхания, но и для опоры. Мангровые заросли — очень топкое место, к тому же на этих территориях часто случаются приливы, отливы или разливы рек. Придаточные корни растут от ствола под углом вниз и дополнительно укрепляют растение на субстрате. Это так называемые ходульные корни, удерживающие огромные стволы с листьями высоко над водой. Такие корни имеет, например, филодендрон.
У некоторых фикусов, например у фикуса бенгальского или баньяна, развиваются столбовидные корни. Они формируются на крупных ветвях, растут вертикально вниз, достигают почвы и укореняются в ней. Постепенно они утолщаются, приобретая вид столбов. Эти корни подпирают огромные ветви фикуса, не давая им сломаться под собственной тяжестью. В Индии растет священная баньяновая роща из одного дерева, занимающая площадь около 5000 м2.
Они формируются на крупных ветвях, растут вертикально вниз, достигают почвы и укореняются в ней. Постепенно они утолщаются, приобретая вид столбов. Эти корни подпирают огромные ветви фикуса, не давая им сломаться под собственной тяжестью. В Индии растет священная баньяновая роща из одного дерева, занимающая площадь около 5000 м2.
Корни могут также прикреплять стебель к опоре. Придаточные корни на стебле плюща помогают ему держаться за кору дерева или цепляться за мельчайшие неровности на стенах, позволяя стеблю подниматься высоко вверх. Это так называемые корни- прицепки.
Корни некоторых тропических деревьев имеют досковидную форму. Они растут от основания ствола вдоль поверхности земли, имеют уплощенную по вертикали форму. Эти корни также укрепляют растение в почве, увеличивая площадь опоры ствола.
Закрепление знаний и умений
Ответьте на вопросы.
Каковы основные функции корня?
Какие виды корней вы знаете?
Какие типы корневых систем вы знаете?
Какая корневая система чаще всего встречается у двудольных растений?
Какая корневая система чаще всего встречается у однодольных растений?
Можно ли управлять развитием корневой системы растения?
Для чего это нужно?
Какие растения образуют корнеплоды?
Какова функция клубеньков на корнях бобовых растений?
Какие еще видоизменения корней вы можете назвать?
Творческое задание. Зарисовать различные видоизменения корней: корнеплоды, корневые клубни, бактериальные клубеньки, воздушные корни, дыхательные корни, столбовидные корни, корни-прицепки, досковидные корни. Их изображение можно найти в дополнительной литературе.
Зарисовать различные видоизменения корней: корнеплоды, корневые клубни, бактериальные клубеньки, воздушные корни, дыхательные корни, столбовидные корни, корни-прицепки, досковидные корни. Их изображение можно найти в дополнительной литературе.
Задания для учеников, интересующихся биологией.
Вырастить несколько растений фасоли. У трех из них удалить кончик корня (провести пикировку), а три оставить без изменения. Посадить растения в ящик для рассады, поставить его на подоконник и не забывать поливать. Через 2 недели аккуратно выкопать все растения, промыть корни и сравнить между собой. Какие изменения произошли с корневой системой пикированной рассады? Как вы это объясните? Загербаризировать результат вашего опыта, подписать и отдать учителю для кабинета биологии.
Провести опыт с проростками фасоли или бобов. Взять проросток и при помощи туши нанести на главный корень метки на равном расстоянии друг от друга. Посмотреть на проросток на следующий день. Что произошло с метками? О чем говорит такое изменение? Подготовить доклад о проделанном опыте.
Посмотреть на проросток на следующий день. Что произошло с метками? О чем говорит такое изменение? Подготовить доклад о проделанном опыте.
Корни стержневые и мочковатые. Учебный фильм
Виды корней и типы корневых систем | Биология 6 класс #21 | Инфоурок
Корень (6 класс) — биология, подготовка к ЕГЭ и ОГЭ 2017
Ресурсы:
И. Н. Пономарёва, О.А. Корнилова, В.С. Кучменко Биология : 6 класс : учебник для учащихся общеобразовательных учреждений
Н. Пономарёва, О.А. Корнилова, В.С. Кучменко Биология : 6 класс : учебник для учащихся общеобразовательных учреждений
Серебрякова Т.И., Еленевский А. Г., Гуленкова М. А. и др. Биология. Растения, Бактерии, Грибы, Лишайники. Пробный учебник 6—7 классов средней школы
Н.В. Преображенская Рабочая тетрадь по биологии к учебнику В В. Пасечника «Биология 6 класс. Бактерии, грибы, растения»
В.В. Пасечника. Пособие для учителей общеобразовательных учреждений Уроки биологии. 5—6 классы
Калинина А.А. Поурочные разработки по биологии 6класс
Вахрушев А.А., Родыгина О.А., Ловягин С.Н. Проверочные и контрольные работы к
учебник «Биология», 6-й класс
Биоуроки http://biouroki.ru/material/lab/2.html
Сайт YouTube: https://www.youtube.com /
Хостинг презентаций
— http://ppt4web. ru/nachalnaja-shkola/prezentacija-k-uroku-okruzhajushhego-mira-vo-klasse-chto-takoe-ehkonomika.html
ru/nachalnaja-shkola/prezentacija-k-uroku-okruzhajushhego-mira-vo-klasse-chto-takoe-ehkonomika.html
Распределение цинка, меди и кадмия в корневой системе фасоли обыкновенной в условиях разных сред произрастания Текст научной статьи по специальности «Биологические науки»
УДК 581.522.5
РАСПРЕДЕЛЕНИЕ ЦИНКА, МЕДИ И КАДМИЯ В КОРНЕВОЙ СИСТЕМЕ ФАСОЛИ ОБЫКНОВЕННОЙ В УСЛОВИЯХ РАЗНЫХ СРЕД ПРОИЗРАСТАНИЯ
© 2010 Ю.В. Макарова1
С помощью гистохимического дитизонового метода установлены закономерности поглощения, локализации и транслокации ионов Zn, Cu и Cd в тканях разных морфологических типов корней у фасоли обыкновенной (Phaseolus vulgaris L.), исследованы концентрационные и средовые зависимости перечисленных процессов.
Ключевые слова: Phaseolus vulgaris L. , корневая система, растительные ткани, гистохимический дитизоновый метод, Zn, Cu, Cd, концентрация, среда произрастания.
, корневая система, растительные ткани, гистохимический дитизоновый метод, Zn, Cu, Cd, концентрация, среда произрастания.
Введение
К настоящему времени в рамках экологической геохимии собран обширный материал, касающийся аккумуляции химических элементов группы тяжелых металлов в пищевых сельскохозяйственных растениях. Неугасающий исследовательский интерес к этой теме обусловлен практической стороной вопроса: именно растениеводческая продукция является начальным звеном в трофической цепи, по которой потенциально опасные элементы могут поступать в пищевой рацион человека и в конечном итоге определять качество его жизни. В абсолютном большинстве случаев поставленная задача решается на основе физических, химических и физико-химических аналитических методов, которые при неоспоримых достоинствах (высокий уровень избирательности, чувствительность, точность) тем не менее не могут ответить на многие частные вопросы, значимые в плане раскрытия механизмов аккумуляции и детоксикации соединений тяжелых металлов.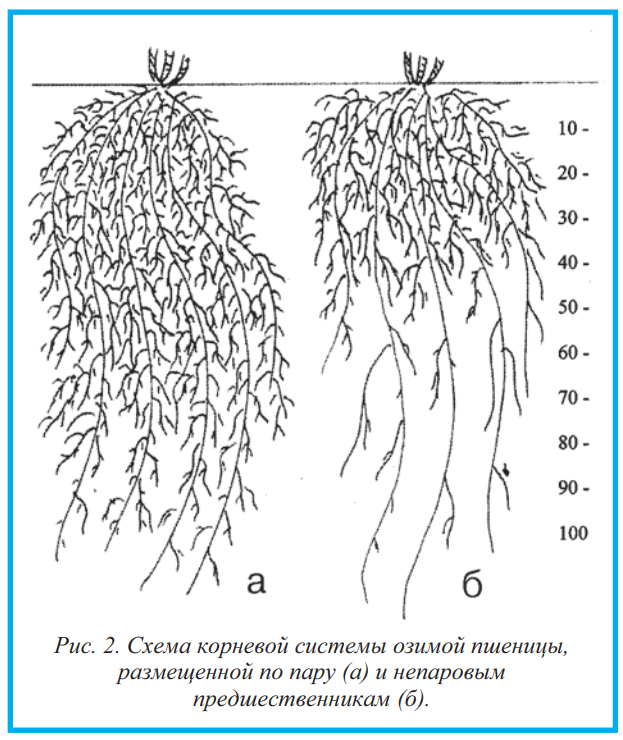 Например, каким образом металлоионы распределяются по органам, тканям и клеткам растений, какова функциональная роль различных тканей в процессах поглощения и транслокации, насколько зависим характер распределения тяжелых металлов в растении от их концентрации в среде произрастания и, наконец, каково влияние субстрата на этот процесс. Справиться с этими вопросами, дополнить и конкретизировать результаты аналитических методов возможно средствами гистохимии. В связи с этим целью настоящих исследований являлось гистохимическое изучение особенностей поглощения, аккумуляции и транслокации в корневой системе
Например, каким образом металлоионы распределяются по органам, тканям и клеткам растений, какова функциональная роль различных тканей в процессах поглощения и транслокации, насколько зависим характер распределения тяжелых металлов в растении от их концентрации в среде произрастания и, наконец, каково влияние субстрата на этот процесс. Справиться с этими вопросами, дополнить и конкретизировать результаты аналитических методов возможно средствами гистохимии. В связи с этим целью настоящих исследований являлось гистохимическое изучение особенностей поглощения, аккумуляции и транслокации в корневой системе
1Макарова Юлия Владимировна ([email protected]), кафедра экологии, ботаники и охраны природы Самарского государственного университета, 443011, Российская Федерация, г. Самара, ул. Акад. Павлова, 1.
фасоли обыкновенной (Phaseolus vulgaris L.), активно вовлекаемых в биологический круговорот Zn, Cu и Cd в зависимости от условий произрастания растения в модельном эксперименте.
Условия и методы исследования
В качестве объектов исследования использовали 14-дневные растения фасоли обыкновенной сорта «Щедрая». Действующим фактором являлись растворы солей ZnSO4-7h3O, CuSO4-3h3O, 3CdSO4-8h3O, которые готовили в пересчете на содержание металла в одном литре. В итоге развитие водной и субстратных культур растения [1] происходило на следующих экспериментальных концентрациях металлов: Zn — 1000,0; 100,0; 10,0 и 1,0 мг/л, Cu — 500,0; 50,0; 5,0 и 0,5 мг/л, Cd — 50,00; 5,00; 0,50 и 0,05 мг/л. В подборе концентраций руководствовались литературными данными [2], согласно которым предельно допустимая концентрация для Zn составляет 1000,0 мг/л, Cu — 500,0 мг/л, Cd — 50,0 мг/л. Обработку субстрата растворами металлов проводили единожды — в момент высадки семян. В остальное время по мере подсыхания его увлажняли дистиллированной водой. В контрольных вариантах для высадки растений (водная культура) и обработки субстрата использовали дистиллированную воду.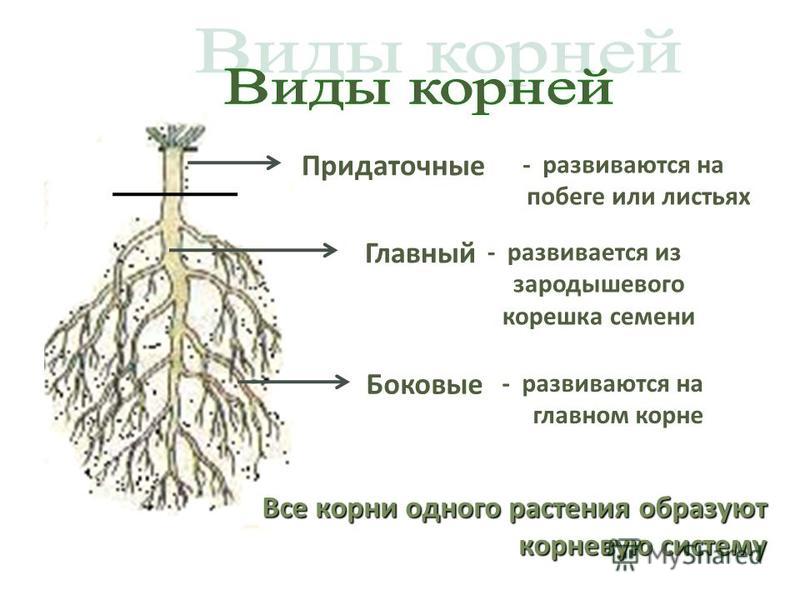
Субстратами для растений служили промытый, прокаленный в муфельной печи речной песок и среднесуглинистый обыкновенный чернозем с содержанием гумусовых веществ 5,9 %, актуальной кислотностью 6,7 ед. и карбонатностью 1,0-2,5 %, отобранный с фонового участка Самарской области [3]. Указанные величины ведущих почвенных характеристик являются вполне благоприятными для роста и развития анализируемого вида [4].
В целях изучения поглощения, аккумуляции и транслокации Zn, Cu и Cd в органах и тканях растения использовали гистохимический дитизоновый метод [5], чувствительность которого в отношении перечисленных ионов составляет 10~5 М/л. Из-за невозможности хранения реактив, состоящий из 3 мг дитизо-на, 6 мл ацетона, 2 мл дистиллированной воды и 1-2 капель ледяной уксусной кислоты, готовили непосредственно перед началом работы. Сделанные при помощи острой бритвы прижизненные срезы главного корня (базальный участок зоны ветвления) и отходящих от него здесь же боковых корней (базальные участки) обрабатывали дитизоновым реактивом и исследовали под световым микроскопом при сменном увеличении до 400 раз, отмечая локализацию и интенсивность окраски структурных элементов по балльной шкале (0 — отсутствие окрашивания, 1 — очень слабое окрашивание, 2 — слабое окрашивание, 3 — окрашивание средней интенсивности, 4 — окрашивание высокой интенсивности, 5 — окрашивание очень высокой интенсивности).
Результаты исследования и их обсуждение
Качественная окраска срезов свидетельствует о присутствии Zn, Cu и Cd во всех тканях корневой системы фасоли независимо от их содержания в средах произрастания. Не являются исключением и контрольные варианты, что может быть обусловлено как первоначальным включением анализируемых химических элементов в структуры покоящегося семени растения, так и процессом их поглощения непосредственно из сред произрастания — песка и почвы (см. рисунок).
Содержание элементов в тканях корневой системы, как правило, определяется их концентрацией в субстрате. При этом корни растений, развивающихся на водных растворах металлов, в силу большей доступности для поглощения аккумулируют Zn, Си и Cd активнее, чем на песке и почве. Экспериментальной особенностью является интенсификация процесса поглощения и, соответственно, накопления химических элементов в корневой системе в вариантах с внесением в субстрат минимальных концентраций металлов (1,0 мг/л Zn, 0,5 мг/л Си, 0,05 мг/л Cd) по сравнению с вариантами, где концентрация металлов в субстрате была на порядок выше (10,0 мг/л Zn, 5,0 мг/л Си, 0,5 мг/л Cd). Выявленная тенденция оказалась несвойственной только для растений, вегетирующих на водных растворах Cd, у которых содержание элемента в тканях главного и боковых корней увеличивается пропорционально росту его концентрации в растворе (см. рисунок). Таким образом, полученные нами данные иллюстрируют не только многократно отмеченное разными авторами и обусловленное возрастанием сосущей силы корневой системы явление усиления поступления ионов металлов в растение при синхронном росте их концентрации в среде произрастания, но и возможность поглощения ионов корневой системой с наибольшей скоростью в случае низких их концентраций в субстрате [6].
60
ВодаПесокрочв; Контроль
ВодаПесоКПочв; 0.05 мг/л Сс1
Вода ПесоКПочваВодаресоКПочва 0.50 мг/л СС 5.00 мг/л СС
Вода ПесокПоч ва 50.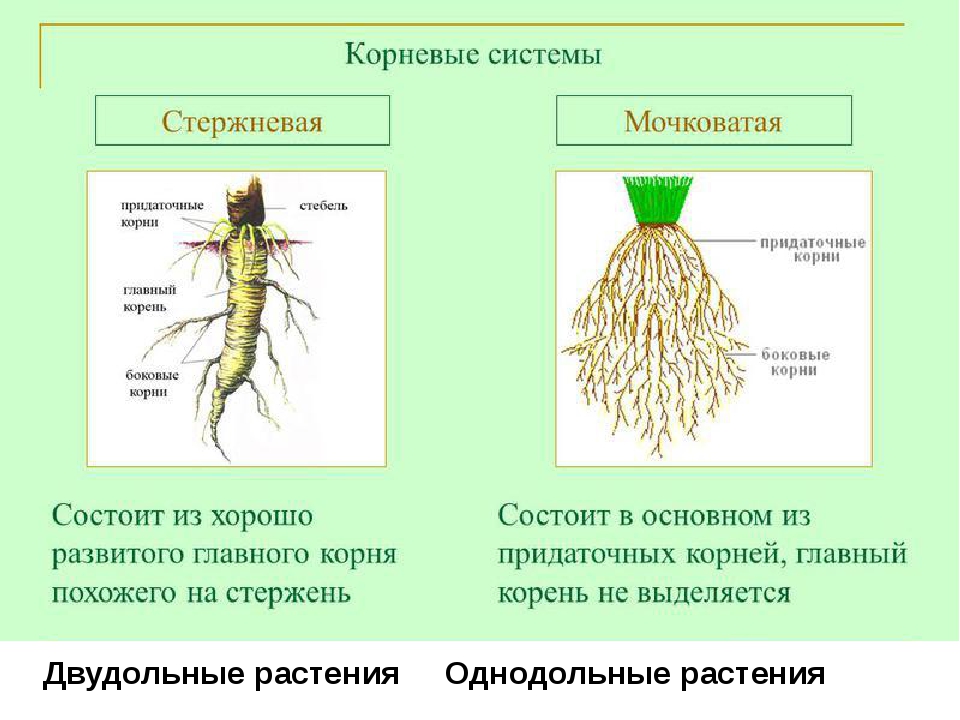 00 мг/л СС
00 мг/л СС
■ Ризодерма ЭЭндодерма
■ Первичная флоэма Ш Пучковый камбий 0 Первичная ксилема
0 Экзодерма □ Перицикл ЕЗ Вторичная флоэма 0 Межпучковый камбий ЕЭ Протоксилема
0 Паренхима первичной коры Н Основная паренхима Ш Флоэма
□ Вторичная ксилема Ш Метаксилема
Рис. Суммарное содержание и распределение Cd по тканям главного и боковых корней фасоли обыкновенной в условиях разных сред произрастания
Уровень аккумуляции металлов в корнях фасоли, принадлежащих к разным морфологическим типам, неодинаков. Практически во всех вариантах на песчаном и почвенном субстрате, а также на водных растворах Cd содержание анализируемых химических элементов в тканях главного корня выше, чем в тканях боковых корней (см. рисунок). Разница между количественным содержанием металлов в этих органах может быть обусловлена рядом причин [7-11].
рисунок). Разница между количественным содержанием металлов в этих органах может быть обусловлена рядом причин [7-11].
Во-первых, по сравнению с позже развивающимися боковыми корнями главный корень обладает большей поглощающей поверхностью. С учетом того, что количественное содержание металлов в тканях корня складывается не только за счет их непосредственного поступления из субстрата по радиусу корня, но и за счет вертикального транспорта по паренхиме первичной коры и ксилеме из нижележащих областей, главный корень, обладая хорошо выраженной, обширной паренхимой первичной коры, более высокоорганизованной проводящей системой и, соответственно, более протяженной поверхностью деятельной части, способен поглощать и накапливать в тканях анализируемого участка Zn, Си и Cd количественно больше, чем боковые корни.
Во-вторых, причиной активизации метаболических явлений в тканях главного корня может являться процесс «зарождения» боковых корней, требующий усиленного притока веществ извне.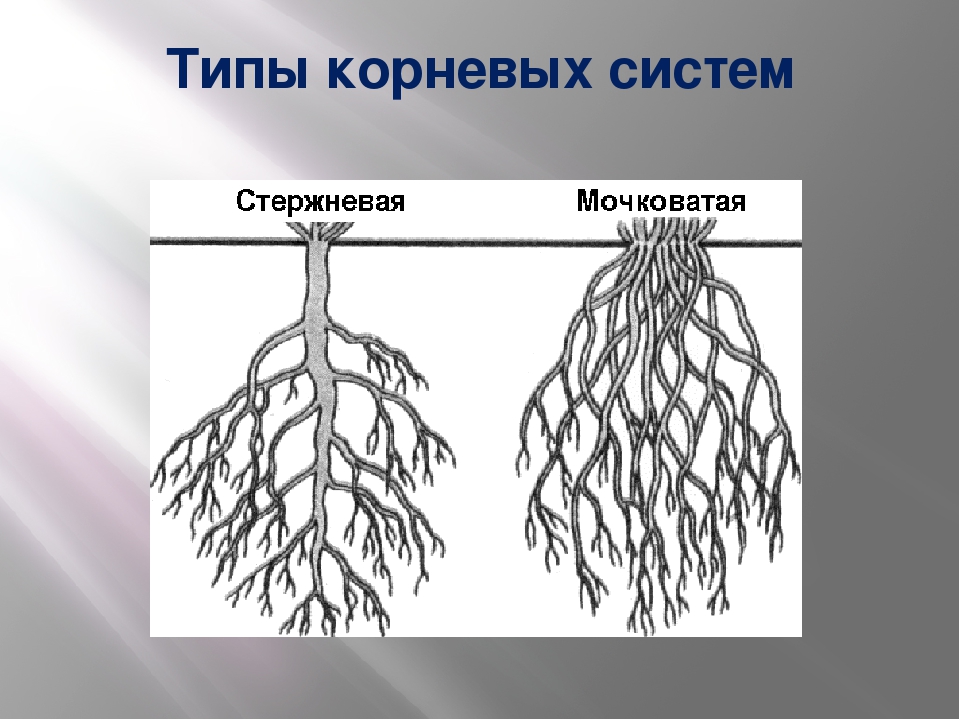 Нами зафиксировано, что рядом с местом разрыва эндодермы примордием бокового корня содержание металлов в тканях центрального цилиндра (вплоть до первичной ксилемы) возрастает, а сам примордий аккумулирует максимально возможное их количество.
Нами зафиксировано, что рядом с местом разрыва эндодермы примордием бокового корня содержание металлов в тканях центрального цилиндра (вплоть до первичной ксилемы) возрастает, а сам примордий аккумулирует максимально возможное их количество.
В-третьих, преобладание металлоионов в главном корне может быть связано с деятельностью боковых корней, которые массово образуются именно на анализируемом участке главного корня, а также характеризуются повышенной метаболической активностью в силу меньшего возраста.
В-четвертых, более развитая проводящая система главного корня обеспечивает возможность большего передвижения воды с растворенными в ней веществами к надземным частям растения, способствуя их развитию. Увеличение надземной фитомассы приводит к возрастанию сосущей силы, которая, действуя на главный корень, ускоряет транспорт веществ от поверхностных тканей к сосудам ксилемы. В свою очередь, форсированное освобождение поглощающей поверхности главного корня будет неизменно приводить к интенсификации его деятельности.
На водных растворах (кроме Cd) и в вариантах с внесением в почву 0,5 мг/л Си и 1 мг/л Zn поглотительная активность главного и боковых корней фасоли практически сходна. Такое распределение металлов между морфологически разными типами корней обусловлено, с одной стороны, их низким содержанием в субстрате, физико-химические и микробиологические характеристики которого (почва) могут дополнительно снижать растворимость соединений и доступность составных элементов для поглощения растению, а с другой — замедлением темпов роста и развития надземной фитомассы вследствие разобщения вещественно-энергетических связей с корневой системой, происходящим на фоне последовательно возрастающих концентраций металлов в водной среде [12].
Гистохимический анализ поперечных срезов корневой системы выявил в ней ряд тканей, поглощающих и накапливающих ионы Zn, Си и Cd наиболее активно. К тканям приоритетного улавливания металлов в главном корне относятся экзодерма, пучковый камбий, ксилема, в боковых корнях — ризодерма, экзодерма, ксилема (см. рисунок).
рисунок).
Причины преимущественного накопления анализируемых металлов в клетках ризодермы вполне очевидны: именно эта ткань является первой на пути их проникновения в корневую систему растения, и именно она, по сравнению с остальными тканями, обладает более развитой и более активной системой мембранных транспортных механизмов [13]. Непосредственный контакт тканей друг с другом обуславливает перемещение ионов Zn, Си и Cd из ризодермы в экзодерму практически в неизменном количестве.
Высокая поглотительная способность пучкового камбия связана не только с интенсивно протекающими в его клетках метаболическими процессами, вызванными возрастной дифференциацией тканей, но и с его близостью к проводящим тканям растения, особенно к ксилеме, обладающей мощной аттрагирующей способностью. Значительное содержание металлов в структурах последней (клетки паренхимной обкладки, стенки сосудов, пасока) может быть предопределено расположением анализируемого участка у основания корня, «запас» элементов в котором создается благодаря их разнонаправленному транспорту. и Cd сконцентрированы, главным образом, в клеточных стенках. На преимущественное связывание металлов клеточными стенками указывают результаты не только гистохимического дитизонового метода [5; 14-16], но и родизонатного метода [17], рентгенфлуорисцентного анализа [18].
и Cd сконцентрированы, главным образом, в клеточных стенках. На преимущественное связывание металлов клеточными стенками указывают результаты не только гистохимического дитизонового метода [5; 14-16], но и родизонатного метода [17], рентгенфлуорисцентного анализа [18].
Протопласты клеток эндодермы и тканей центрального цилиндра (особенно паренхимы, расположенной вблизи крупных сосудов ксилемы), как правило, накапливают анализируемые металлы в большем количестве, чем клетки коровой паренхимы. Содержание химических элементов в протопластах, а также в стенках сосудов и пасоке ксилемы находится в прямо пропорциональной зависимости от их концентрации в субстрате.
Помимо существующих между вариантами различий в характере распределения и в уровне аккумуляции металлов в тканях на песчаном (0,05-50,00 мг/л Cd) и почвенном (5,00 и 50,00 мг/л Cd) субстратах в срединной области первичной коры главного корня нами зафиксировано отмирание отдельных клеток или значительно чаще — их групп. Возникающие полости пространственно разделены между собой живыми паренхимными клетками, организованными в радиально ориентированные тяжи.
Возникающие полости пространственно разделены между собой живыми паренхимными клетками, организованными в радиально ориентированные тяжи.
В качестве главной причины этого явления классически называют рост корней в условиях затрудненной аэрации, когда образуемые растением полости служат резервуарами необходимого для дыхания тканей кислорода [19]. Но в связи с тем, что в эксперименте развитие полостей происходит только в вариантах с внесением в песчаный и почвенный субстрат одного элемента — Cd, с большей долей вероятности можно говорить об их образовании как о приспособительной реакции растения на действие высокотоксичного вещества [20]. Накапливаясь во внут-рикорневых полостях и диффундируя из них в прикорневую область субстрата, молекулы кислорода могут принимать участие в реакциях окисления соединения Cd, снижая его негативное воздействие на рост и развитие растительного организма.
«Уникальность» действия Cd, широта диапазона его концентраций в условиях разных сред произрастания свидетельствуют, что на процесс образования полостей в структуре паренхимы первичной коры влияют не только химическая природа элемента и его концентрация в среде, но и физико-химические характеристики субстрата, в частности, обусловленная присутствием глинистых минералов и органического вещества его обменная емкость. локализованы в симпласте тканей центрального цилиндра и в апопласте внешнего по отношению к нему кольца тканей. В случае значительного содержания металлов в среде произрастания их активное поступление в центральный цилиндр происходит и по апопласту, и по симпласту.
локализованы в симпласте тканей центрального цилиндра и в апопласте внешнего по отношению к нему кольца тканей. В случае значительного содержания металлов в среде произрастания их активное поступление в центральный цилиндр происходит и по апопласту, и по симпласту.
Литература
[1] Практикум по агрохимии / под ред. В.Г. Минеева. М.: Изд-во МГУ, 2001. 689 с.
[2] Вредные химические вещества. Неорганические соединения элементов I-IV групп / под ред. В.А. Филонова. Л.: Химия, 1988. 512 с.
[3] Прохорова Н.В., Матвеев Н.М., Павловский В.А. Аккумуляция тяжелых металлов дикорастущими и культурными растениями в лесостепном и степном Поволжье. Самара: Изд-во «Самарский университет», 1998. 131с.
[4] Лебедева Г.Ф. Пособие по растениеводству для почвоведов.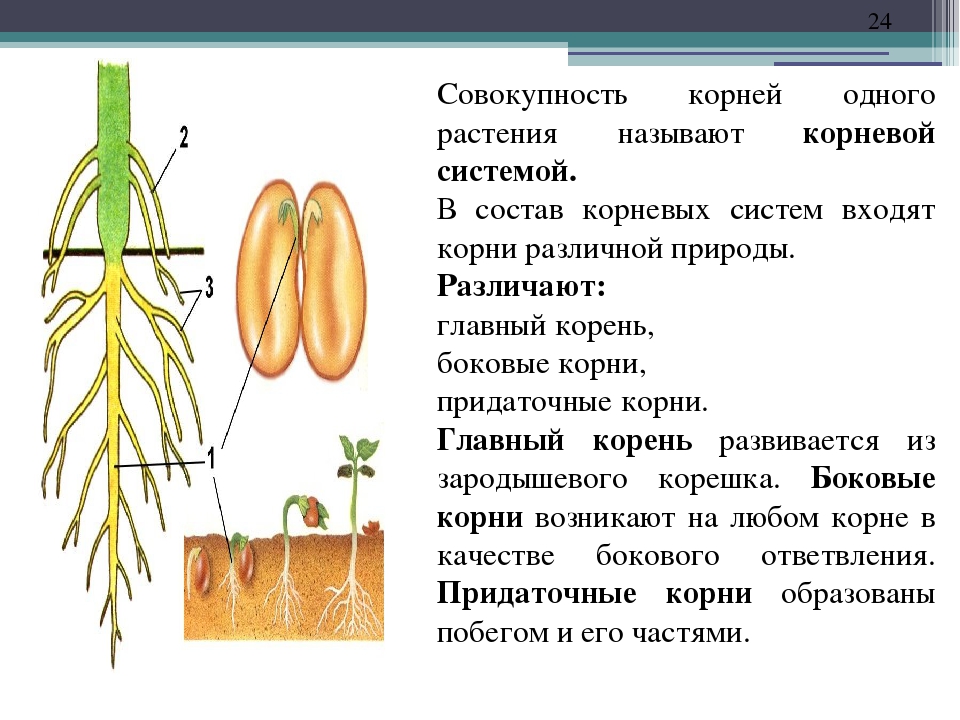 Зерновые и зерновые бобовые культуры: в 2 ч. М.: Изд-во МГУ, 1982. Ч. 2. 137 с.
Зерновые и зерновые бобовые культуры: в 2 ч. М.: Изд-во МГУ, 1982. Ч. 2. 137 с.
[5] Серегин И.В., Иванов В.Б. Передвижение ионов кадмия и свинца по тканям корня // Физиология растений. 1998. Т. 45. № 6. С. 899-905.
[6] Воробьев Л.Н. Регулирование мембранного транспорта в растениях // Итоги науки и техники. Физиология растений. 1980. Т. 4. С. 1-77.
[7] Колосов И.И. Поглотительная деятельность корневых систем растений. М.: Изд-во АН СССР, 1962. 338 с.
[8] Сытник К.М., Книга М.Н., Мусатенко Л.И. Физиология корня. Киев: Наукова думка, 1972. 356 с.
9] Нестерова А.Н. Действие тяжелых металлов на корни растений. 1. Поступление свинца, кадмия, цинка в корни, локализация металлов и механизмы устойчивости растений // Биологические науки. 1989. № 9. С. 72-86.
1989. № 9. С. 72-86.
10] Трапезников В.К., Иванов И.И., Тальвинская Н.Г. Локальное питание растений. Уфа: Гилем, 1999. 260 с.
11] Физиология растений / под ред. И.П. Ермакова. М.: Издательский центр «Академия», 2007. 640 с.
12] Макарова Ю. В. Изменчивость морфометрических показателей фасоли обыкновенной при краткосрочном воздействии кадмия, цинка и меди // Вестник СамГУ. Естественнонаучная серия. 2009. № 6 (72). С. 160-171.
13] Вахмистров Д.Б. Пространственная организация ионного транспорта в корне // 49-е Тимирязевские чтения. М.: Наука, 1991. С. 48.
14] Серегин И.В., Иванов В.Б. Физиологические аспекты токсического действия кадмия и свинца на высшие растения // Физиология растений. 2001. Т. 48. № 4. С. 606-630.
15] Башмаков Д.И. Аккумуляция тяжелых металлов некоторыми высшими растениями в разных условиях местообитания // Агрохимия. 2002. № 9. С. 66-71.
16] Кожевникова А.Д. Распределение никеля, кадмия, свинца и стронция в прорастающих зерновках кукурузы // VIII молодеж. конф. ботаников: тез. докл. СПб.: СПбГУТД, 2004. С. 127-128.
17] Серегин И.В. Кожевникова А.Д. Транспорт, распределение и токсическое действие стронция на рост проростков кукурузы // Физиология растений. 2004. Т. 51. № 2. С. 241-248.
18] Wierzbiska M., Antosiewicz D. How lead can easily enter the food chain a study of plant roots // Sci. Total Environ. Suppl. 1993. № 1. P. 423-429.
19] Березина Н.А., Афанасьева Н.Б. Экология растений. М.: Издательский центр «Академия», 2009. 400 с.
20] Эзау К. Анатомия семенных растений: в 2 кн. М.: Мир, 1980. 558 с.
Поступила в редакцию 15/VT/2010; в окончательном варианте — 15/VT/2010.
CONCERNING THE DISTRIBUTION OF ZINC, COPPER AND CADMIUM IN THE ROOT SYSTEM OF KIDNEY BEAN UNDER DIFFERENT GROWTH MEDIA
© 2010 Ju.V. Makarova2
Using the histochemical dithizonic method the patterns of absorption, localization and translocation of Zn, Cu and Cd ions were studied in different morphological tissues of the kidney bean (Phaseolus vulgaris L.) roots. These processes were also investigated in connection with the concentration and media composition parameters.
Key words: Phaseolus vulgaris L., root system, plant tissues, histochemical dithizonic method, Zn, Cu, Cd, concentration, growth media.
Paper received 15/VI/2010. Paper accepted 15/VI/2010.
2Makarova Julia Vladimirovna (aconithumamail.ru), the Dept. of Ecology, Botany and Environmental Protection, Samara State University, Samara, 443011, Russian Federation.
Тема: КОРЕНЬ | Лабораторные работы по биологии
Тема: КОРЕНЬ
25.09.2014 6501 0Лабораторная работа 9. Типы корневых систем. Строение стержневой и мочковатой корневых систем
Цели: познакомить учащихся со строением и развитием стержневых и мочковатых корневых систем.
Оборудование: проростки семян фасоли (гороха, огурца и т.д.), зерновок пшеницы (ржи, кукурузы и т.д.), в возрасте, 5-7 дней и 10-12 дней; гербарии растений со стержневой и мочковатой корневыми системами.
Ход работы:
1. Рассмотрите проросшие семена фасоли 5-7 дневного возраста. Какой корень вы видите?
2. Рассмотрите проросшие семена фасоли 10-12 дневного возраста. Появились ли какие-нибудь изменения в корневой системе?
3. Рассмотрите корневую систему взрослого растения фасоли. Найдите главный и боковые корни. Как называется такой тип корневой системы?
4. Зарисуйте корневую систему проростка фасоли в возрасте 5-7 дней и 10-12 дней, а так же корневую систему взрослого растения фасоли. Подпишите тип корневой системы и все виды корней.
5. Рассмотрите проросшие зерновки пшеницы 5-7 дневного возраста. Сколько корней появляется на первой стадии развития проростка пшеницы?
6. Рассмотрите проросшие зерновки пшеницы 10-12 дневного возраста. Как происходит формирование корневой системы пшеницы? Какие типы корней вы видите? В чем отличие развития корневой системы пшеницы от корневой системы фасоли?
7. Рассмотрите корневую систему взрослого растения пшеницы. Можете ли вы различить главный корень? Найдите придаточные и боковые корни.
8. Зарисуйте корневую систему проростка пшеницы в возрасте 5-7 дней и 10-12 дней, а так же корневую систему взрослого растения пшеницы. Подпишите тип корневой системы и все виды корней.
9. Сравните корневые системы фасоли и пшеницы.
10. Сделайте вывод о сходствах и различиях корневых систем фасоли пшеницы.
Ботаническая характеристика фасоли
Растения фасоли самоопыляющиеся, но в жару, на юге, опыляются и перекрёстно, преимущественно насекомыми. Растения фасоли имеют кустовую или полу кустовую форму с небольшими боковыми побегами, дающими цветки и бобы, а также вьющуюся форму, которая образует побеги с длинными междоузлиями.
Стебель травянистый, иногда деревенеющий у основания, зелёной, серо-зелёной, розовой или фиолетовой окраски. Высота стебля кустовых форм 20…60 см, вьющихся — до 2…5 метров.
Листья светло-зелёные или темно-зелёные, у сортов с тёмноокрашенными семенами черешки фиолетовые. Листья непарноперистые, состоящие из трёх листовых пластинок на общем черешке. Их величина, форма и окраска изменяются в зависимости от сорта и условий выращивания. Все надземные вегетативные органы покрыты опушением.
Корневая система овощной фасоли развита слабо и расположена в поверхностном слое почвы, только отдельные корни проникают на глубину 1 м. На корнях образуется множество клубеньков, в которых поселяются бактерии, усваивающие свободный азот из воздуха.
Цветки мелкие, белые, розовые, фиолетовые самых различных оттенков. Они расположены на коротких цветоносах попарно. Между окраской лепестков и семян существует связь — у сортов с более темными лепестками семена более тёмные. Цветение фасоли начинается на 40…60-й день после посева и продолжается 15…20 дней. Через две-три недели после оплодотворения развиваются плоды.
Бобы завязываются в пазухах листьев. В фазе технической спелости они окрашены в желто-восковой (у спаржевой фасоли), светло- или темно-зеленый цвет, иногда со светло-фиолетовой штриховкой. У лущильных сортов на внутренней стороне створки имеется грубый толстый пергаментный слой; у полусахарных — слабый и поздноразвивающийся слой с волокнами по краям; у сахарных нет ни волокон, ни пергаментного слоя. Семена одноцветные или пёстрые, чаще белые. Их в бобе 4…10 шт. Масса 1000 семян 150…500 г. Старые семена теряют блеск. Всхожесть сохраняется в течение 4…5 лет и более.
Следует сказать, что у диких форм фасоли белых семян не было. Белозерные сорта выведены селекционерами и более пригодны для приготовления супов и консервов, поскольку дают светлый прозрачный бульон.
Вьющиеся формы возделывают в основном на индивидуальных огородах и приусадебных участках.
Виды корневых систем
У наземных цветковых растений над землей находится побеги с зелеными листьями, а в почве — корни. Корень — это орган, обеспечивающий растение водой и минеральными веществами и укрепляющий его в почве. Корни, всасывая из почвы с растворенными в ней минеральными веществами, обеспечивают почвенное питание растений. С помощью корней растения прочно укрепляются в почве. Это легко доказать. Попробуйте вырвать с корнями осот, бодяк или другой крупный сорняк. Оказывается это не так-то и легко сделать! В корнях образуются многие важные для жизни растений растений вещества, в том числе некоторые витамины. В корнях моркови, сельдерея и некоторых других растений могут накапливаться запасные вещества.
Каждое
растение имеет множество корней. Развитие
корней можно проследить в опытах с
проращиванием семян в приборе для
наблюдения за развитием корневой системы у
растений.
При проращивании семян фасоли или гороха
через 3-4 дня после замачивания разрывается
семенная кожура и появляется корешок
зародыша. Он быстро растет и со временем
развивается в главный корень растения. У
многих двудольных растений, в том числе и у
фасоли, и у гороха, главный корень растет
отвесно вниз и выделяется среди прочих
корней большей длиной и толщиной. Такой
корень похож на стержень. Через 1-2 дня после
начала прорастания на главном корне
появляются боковые корни. Они развиваются в
строгой последовательность. Первые из них
появляются вблизи основания главного корня,
а последующие по мере его роста — ближе к
кончику. Наиболее старые и соответственно
самые длинные боковые корни находятся
поэтому вблизи поверхности почвы, а более
молодые и короткие — на большой глубине.
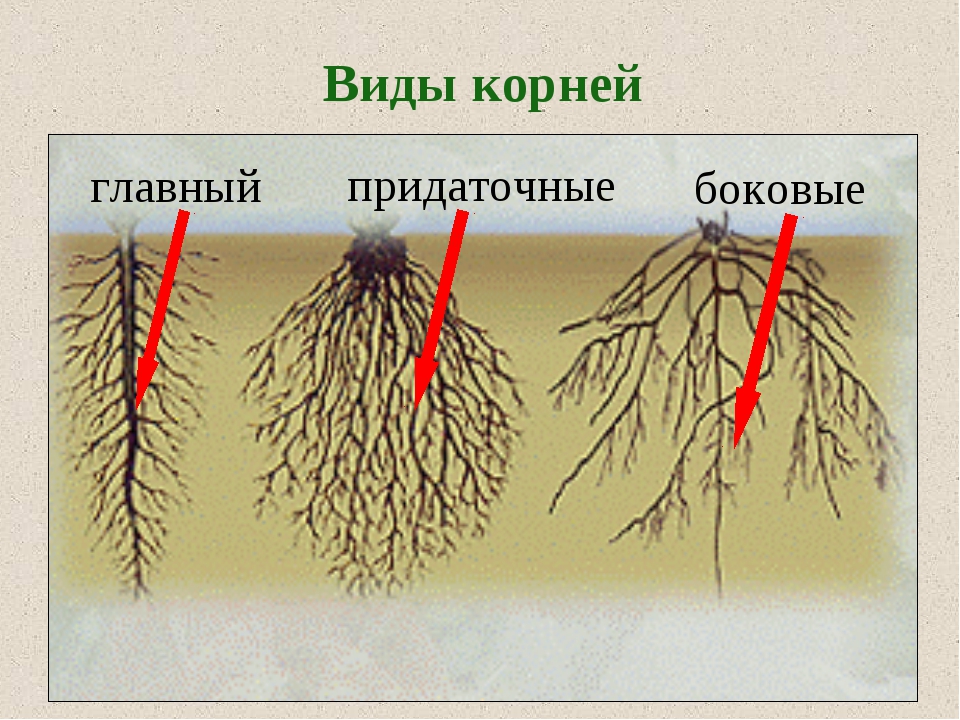
У фасоли и гороха корни появляются также на нижней части побега. Граница между стеблем и главным корнем у фасоли и гороха четко видна, поскольку стебель немного толще корня. Вырастающие на побегах корни называют придаточными. Придаточные корни в отличие от боковых образуются без строгой очередности, и длина их поэтому колеблется беспорядочно. Чтобы усилить появление придаточных корней, растение окучивают, подсыпая к основаниям побегов землю. Окучивание повышает урожай таких растений, как картофель, томат, кукуруза, и некоторых других сельскохозяйственных растений.
Корневые системы
Боковые и придаточные корни ветвятся. В итоге одно растение развивает множество корней, общая длина которых достигает несколько сотен и даже тысяч метров. Совокупность всех корней одного растения составляет его корневую систему.
Корни. Фото:
Tim Green
Корневые системы, в которых можно различить главный корень, называют стержневыми. Такие корневые системы характерны для фасоли, гороха, свеклы, подсолнечника и многих других двудольных растений.
У однодольных растений, например лука, пшеницы, зародышевый корешок быстро прекращает рост. От оснований побегов у них отрастают многочисленные придаточные корни. Растение развивает пучок или мочку придаточных корней, более-менее одинаковых по толщине, длине и разветвленности.
Корневые системы, образованные множеством обильно ветвящихся придаточных корней, называют мочковатыми. Такие корневые системы свойственны хлебным злакам и другим однодольным растениям. Встречаются они и у некоторых двудольных растений, например у лютика, подорожника, первоцвета.
Корневые системы взрослых растений занимают большой объем почвы. У пшеницы и ржи, например корни распространяются вширь на 1-1,5 м и проникают вглубь до 2 м, при общей длине 500-600 м. Размеры корневых систем обычно намного больше, чем размеры надземных органов. Так, у люцерны длина главного корня в 10 раз превосходит высоту надземной части растения. У плодовых деревьев диаметр корневых систем в 2-5 раз больше диаметра кроны. Изучать корневые системы очень трудно, так как они скрыты в земле. Их изучение требует или тщательных и трудоемких раскопок, или использования специального оборудования: корнеотмывочных машин, с раздвижными стеклянными стенками. По этим причинам мы пока знаем о строении корневых систем намного меньше, чем о строении надземных органов.
В
течение жизни растение постоянно
увеличивает размеры своих корней. При этом
они углубляются в почву и разрастаются
далеко в стороны от стебля. Корни свеклы,
например, проникают в почву на глубину
более 3 м, у кукурузы – 2–2,5 метра, у люцерны
– до 5 метров, у пшеницы – на 2-2,5 м. У огурцов
корни разрастаются на 1,5-2 м в
стороны от стебля, у лука – на 60-70 см. У
взрослой яблони корни разрастаются в
стороны на 10-12 м от ствола, а у осины – даже
на 30 м.
Корни обладают неограниченной
возможностью роста. Но в природе они редко
реализуют такую возможность из-за
различных причин: ветвления в почве корней
других растений, недостаточности
питательных веществ и т. д. В специально
созданных условиях растение способно
развить огромную массу корней. Например, у
четырехмесячного растения озимой ржи,
выращенного в теплице, образовалась
огромная корневая система. Общая длина всех
ее корней составила 623 км, что равно
расстоянию от Москвы до Санкт-Петербурга.
Причем общая поверхность всех корней этого
растения была в 130 раз больше поверхности
наземных органов растения. Общий прирост
всех корней только за одни сутки достигал 4,8
км, и ежесуточно на них образовывалось
более 100 млн корневых волосков.
Корни растут в течение всей жизни растения.
Рост
корня осуществляется путем деления и
растяжения клеток, находящихся на верхушке
(кончике) корня. Все корни растут
верхушечной частью. Обнаружить это можно с
помощью следующего опыта. На корешок
проростка фасоли или тыквы нанесите черной
тушью тонкие черточки-метки на одинаковом
расстоянии друг от друга. Уже через день
можно увидеть, что на участке,
расположенном ближе к верхушке корня,
расстояние между метками увеличилось,
тогда как у основания корня оно не
изменилось.
Если удалить верхушку, то рост корня в длину
прекратится, но зато образуется много
боковых корней. Эта особенность растений
используется человеком при выращивании
рассады культурных растений. Корень
удлиняется и продвигается в почве в
результате деления и растяжения клеток
верхушки корня.
Типы корневой системы зависят от ее происхождения. При семенном размножении растений корневая система характеризуется наличием явно выраженного стержневого корня; при черенковом — имеет достаточно развитые вертикальные и горизонтальные корни при отсутствии стержневого. Отпрысковая корневая система характеризуется преимущественным развитием горизонтальных корней и небольшой глубиной их залегания.
Корневые системы семенного происхождения имеют сеянцы растений, а также культурные плодовые деревья с привитыми на них сортами. Они имеют главный (стержневой) корень и много боковых, которые с возрастом превращаются в горизонтальные и вертикальные, образуя разветвления корней разных порядков. У поверхности почвы растения, имеющие такую корневую систему, имеют расширенную часть отвода, так называемую корневую шейку — место перехода надземной части ствола в подземную. При перенесении саженца на постоянное место в сад важно не заглубить дерево ниже корневой шейки, т.е. посадить на такую глубину, на какой саженец рос в питомнике. При значительном заглублении деревья плохо растут, имеют метельчатую форму, мало плодоносят и не имеют производственного значения.
Корневая
система семенных растений образует
значительное количество вертикальных
корней и проникает в почвогрунт до глубины
3-10 м и более в зависимости от вида и
разновидности подвоя, а также качества
почвы и глубины залегания грунтовых вод.
Это необходимо учитывать при закладке сада
на семенных подвоях.
Корневые системы, возникающие из черенков и
отводков, при вегетативном способе
размножения наиболее характерны для таких
плодовых растений, как дусен, парадизка,
айва, а также корнесобстственных сортов
плодовых деревьев. Они имеют достаточно
развитые вертикальные, и особенно
горизонтальные корни при отсутствии
главного (стержневого). Корни проникают на
глубину до 2-3 м. Саженцы не имеют корневой
шейки, но место перехода надземной части в
подземную называют условной корневой
шейкой, которую легко можно определить по
цвету коры. У припочвенной части ствол
зеленовато-серый, а у заглубленной -
красновато-коричневый. Саженцы, привитые на
вегетативно размножаемых подвоях,
необходимо садить до условной корневой
шейки.
Корневые системы отпрысковых растений характерны для деревьев, обладающих способностью образовывать побеги из спящих (адвентивных) почек на своих корнях. Из таких побегов вырастают новые плодовые деревья. Корни располагаются в почвогрунте главным образом горизонтально и на небольшой глубине (20-40 см), а незначительная часть их проникает вглубь до 2-3 м. Из плодовых деревьев такую корневую систему имеют корнеотпрысковые вишни, сливы, терн. Настоящей корневой шейки они не имеют, а условная расположена у поверхности почвы. Сажать деревца или сорта, привитые на них, необходимо только до условной корневой шейки, которую хорошо видно на выкопанном отпрыске. В нижней части стволик зеленый или серовато-зеленый, а в заглублении - темно-коричневый.
Описанные типы корневых систем характеризуются большим разнообразием форм. Та или иная конкретная форма корневой системы имеет значение в определении густоты посева семян и посадки растений, глубины обработки почвы, глубины внесения удобрений. Исследование форм корневой системы позволяет судить о взаимодействии ее с почвогрунтом в конкретных почвенно-климатических условиях.
Изученные автором корневые системы подвоев для яблони, груши, сливы, черешки, вишни, абрикоса и персика имели большое разнообразие форм. В одних случаях вертикальные корни были рассредоточены по периферии раскопок, в других — только в срединной части. Большое разнообразие естественных форм корней трудно привести к сопоставимой системе. Поэтому возникает потребность построения такой системы для научных и практических целей. Одним из путей здесь, по нашему мнению, может явиться построение на основе полученных при раскопках количественных данных условных графических изображений корневых систем. За основу нами приняты показатели измерений общей длины всех корней в каждом 20-сантиметровом слое почвогрунте. Большинство изученных нами корневых систем плодовых деревьев можно свести к четырем характерным графическим изображениям, имеющим формы: веретенообразную с сужающимся заостренным хвостом; веретенообразную с широким хвостом; веретенообразную с широким, переменносужающимся хвостом; моркововидную.
Пространственное картирование притока фосфора в корневую систему фасоли с помощью цифровой авторадиографии | Журнал экспериментальной ботаники
Аннотация
Новый метод был разработан для пространственного картирования способности чистого притока фосфора в неповрежденных корневых системах. Метод основан на цифровой авторадиографии и позволяет количественно определять приток фосфора с высоким пространственным разрешением (2 мм). Корни 18-дневных растений фасоли подвергали воздействию 32 Р-меченого ортофосфата, быстро замораживали, вырезали, лиофилизировали, сканировали и подвергали хранению на люминофорном экране.Графики зависимости содержания 32 P от длины корня (расстояние от кончика корня или от основания корня) были получены для трех различных классов корней: базальных, базальных боковых и боковых сторон главного корня. Радиоактивность, обнаруженная методом беспленочной авторадиографии, хорошо коррелировала ( r 2 = 0,99) с измерениями, выполненными сцинтилляционным счетом. Базальные корни абсорбировали фосфора в 2,5 и 1,9 раза больше, чем классы боковых стержневых корней и боковые базальные корни, соответственно, в первые 20 мм от верхушки корня.Внешний фосфор заметно повлиял на приток: корни в среднем составляли 5, 16 и 34 пмоль P мин -1 в апикальных 20 мм при воздействии 1, 5 и 10 мкМ растворов P соответственно. Пространственная картина притока фосфора вдоль корневых осей различных классов корней была довольно однородной при измерении на основе площади поверхности корня. Приток фосфора в более старые сегменты базальных корней (соседние с гипокотилем) не отличался от более новых сегментов, расположенных ближе к вершине корня. Однако при измерении по длине для базальных корней была обнаружена неоднородная картина, что указывает на то, что как класс корня, так и диаметр являются основными факторами, контролирующими пространственную структуру чистого притока.
Введение
Корневые системы состоят из сложного массива различных корневых классов (Fitter, 1991; Lynch, 1995). Различные классы корней используют разные домены почвы, подвержены различным внешним сигналам и могут обладать разной внутренней способностью поглощать питательные вещества. Рассел и Сандерсон (1967) обнаружили большие различия в притоке фосфора между семенными, узловыми и боковыми корнями ячменя. «Специализация» корневого класса по усвоению питательных веществ может иметь практическое значение.Например, большая способность придаточных корней к притоку фосфора будет полезна для растения, поскольку этот класс корней обычно исследует верхний слой почвы, где находится большая часть доступного почвенного фосфора (Pothuluri et al ., 1986; Lynch and Brown, 2001 ; Rubio et al ., 2001).
Пространственная неоднородность притока фосфора также может ожидаться вдоль корневой оси. Такая информация может быть важной при построении моделей роста корней (Lynch and Nielsen, 1996), поглощения питательных веществ (Clarkson, 1996; Silberbush, 1996) и общей функции корней (Aguirrezabal et al ., 1993; Эшель и Вайзель, 1996). Однако современные модели обычно предполагают постоянный характер притока вдоль корневой оси (Smethurst and Comerford, 1993; Rubio et al ., 2001). Большинство классов корней имеют выраженную морфологическую и анатомическую дифференциацию от кончика до основания. Эти секции обычно представляют собой верхушку корня, зону растяжения, зону созревания и зону созревания. Считается, что кончик корня, который включает в себя корневой покров и меристематическую область, имеет большую потребность в питательных веществах (Clarkson, 1996).В зоне созревания развитие корневых волосков из эпидермиса и функциональная лента Каспариана в энтодерме способствуют усвоению питательных веществ. В зрелой области корня кора может стареть, могут появиться боковые корни, а у двудольных растений важен вторичный рост. Степень вакуолизации клеток увеличивается от кончика корня (где нет клеточных вакуолей) к основанию корня. Связь анатомической неоднородности вдоль корневой оси с поступлением питательных веществ в течение некоторого времени беспокоила исследователей (Prevot and Steward, 1936) и, по-видимому, зависит от питательных веществ.Например, поглощение железа, по-видимому, сосредоточено на кончике корня (Clarkson, 1996), а поглощение серы — в зоне растяжения сразу за меристематической областью (Holobrada, 1977). Колмер и Блум (1998) обнаружили стойкие различия в характере поглощения азота корнями кукурузы и риса, вероятно, вызванные наличием слоя склеренхиматозных волокон в корнях риса. В случае фосфора были получены противоположные результаты. Некоторые исследователи отметили, что все участки корня эффективно поглощают фосфор (Burley et al ., 1970), в то время как другие сообщили, что фосфор более активно абсорбируется в апикальных областях корневой оси (Bar-Yosef et al ., 1972).
Для количественной оценки способности поглощения питательных веществ с высоким пространственным разрешением наиболее распространенными подходами были стационарные или вибрирующие ионоселективные микроэлектроды и индикаторы радионуклидов. Ионоселективные стационарные микроэлектроды использовались для определения поступления аммония и нитратов в ячмень (Colmer and Bloom, 1998; Henriksen et al ., 1990) и калия в кукурузе (Newman et al ., 1987). Вибрационные микроэлектроды первоначально использовались для измерения градиентов внеклеточного кальция, а затем использовались для измерения потоков калия и кальция в корнях кукурузы (Kochian et al ., 1992). Микроэлектродные подходы не были полезны для изучения поглощения фосфора из-за отсутствия хороших чувствительных к фосфору электродов. Емкость корня по поглощению фосфора измерялась в основном с помощью меченых ионов (Epstein et al ., 1963; Макнотон и Чапин, 1985; Rubio и др. ., 1997). Стандартные авторадиографические методы заключаются в воздействии на исследуемый объект радиоактивного соединения. Затем объект с радиоактивной меткой подвергается воздействию фотоэмульсии для получения карты распределения радионуклидов (Yeung, 1984). Стандартная авторадиография широко использовалась для изучения динамики и расширения зон истощения питательных веществ (Jungk, 1987; Krauss et al ., 1987; Owusu-Bennoah and Wild, 1979), радиального распределения фосфора в различных областях корней. (Ferguson and Clarkson, 1976), а также скорость поглощения фосфора различными корневыми зонами (Clarkson et al ., 1978). Совсем недавно цифровая авторадиография была использована для количественного описания транспорта гиф 32 P в микоризных культурах корней (Nielsen et al ., 2002) и морфологических характеристик корня, таких как диаметр корня, длина и характер ветвления (Moran et al. ., 2000).
Целью данной работы было оценить изменение притока фосфора вдоль оси у различных классов корней фасоли обыкновенной. Используемый здесь экспериментальный подход включал два этапа.Во-первых, были проанализированы экспериментальные условия, в которых перемещением фосфора вдоль корневой оси можно пренебречь, чтобы можно было оценить чистый приток в каждую корневую секцию. С этими результатами на втором этапе был разработан новый метод количественной авторадиографии для оценки притока фосфора с высоким пространственным разрешением.
Материалы и методы
Растениеводство
Корневая система фасоли обыкновенной ( Phaseolus vulgaris L.) состоит из стержневого корня, придаточных корней и зонтика основных корней, на которых остальная корневая система развивается посредством бокового ветвления (рис. 1). Для корней фасоли обыкновенной, Wanke et al . (1998) сообщили, что расположение различных зон вдоль оси следующее (расстояния от верхушки корня): кончик корня 0–7 мм, зона удлинения 7–20 мм, зона созревания 20–25 мм. Семена фасоли обыкновенной генотипа BAT 477 стерилизовали поверхность в течение 1 мин в 10% NaOCl и проращивали при 25 ° C в течение 2 дней.BAT 477 представляет собой генотип со средним посевом (22 г на 100 семян) и принадлежит к расе М мезоамериканского генофонда (Singh et al ., 1991). Однородные проростки пересаживали в 20-литровые контейнеры, заполненные твердофазным буфером из чистого кварцевого песка (Lynch et al ., 1990), сохраняя либо низкую (1 мкМ), либо высокую (30 мкМ) доступность фосфора. Растения орошали каждые 2 дня питательным раствором без фосфора, состоящим из (в мкМ): 1500 KNO 3 , 1200 Ca (NO 3 ) 2 , 400 NH 4 NO 3 , 25 MgCl 2 , 5 Fe-Na-EDTA, 500 MgSO 4 , 300 К 2 SO 4 , 300 (NH 4 ) 2 SO 4 , 1.5 MnSO 4 , 1,5 ZnSO 4 , 0,5 CuSO 4 , 0,5 Na 2 B 4 O 7 и 0,143 (NH 4 ) 6 Mo 7 O 24 . Чтобы свести к минимуму накопление питательных веществ в питательной среде, во время полива поддерживалась 100% -ная фракция выщелачивания. Растения выращивали в теплице с регулируемой температурой (22–30 ° C) в Юниверсити-парке, штат Пенсильвания (40 ° 85 ′ с.ш., 77 ° 83 ′ з.д.), со средней фотосинтетически активной радиацией в полдень в 1000–1600 мкмоль фотонов на метр –2. с −1 .Общее описание роста и структуры корневой системы проростков фасоли дано в Lynch and van Beem (1993).
Рис. 1.
Схема корневой системы обычного бина, показывающая различные корневые классы.
Рис. 1.
Схема корневой системы обычного боба, показывающая различные корневые классы.
Осевой транспорт фосфора
Поскольку предлагаемый метод подразумевает баланс масс на уровне сегмента корня, сначала было определено время воздействия, при котором транслокацией можно пренебречь.С этой целью был проведен эксперимент, в котором отдельные участки корней подвергались воздействию меченого фосфора, чтобы оценить перемещение фосфора по осям корня и экспериментальные условия, в которых им можно пренебречь.
Через 18 дней роста базальные корни растений фасоли помещали на блоки из оргстекла размером 3 × 18 × 3 см с 12 ячейками, каждая 1,5 см глубиной и 0,5 см шириной (рис. 2). Растения выращивали в условиях с высоким содержанием фосфора, при которых максимальный уровень мобилизации фосфора из корней наблюдается у фасоли обыкновенной (Snapp and Lynch, 1996).Каждая ячейка была соединена со следующей ячейкой неглубоким мостом, так что корень можно было проложить поперек блока, пересекая все ячейки. Все клетки содержали 5 мкМ фосфата калия и 0,5 мМ CaSO 4 для поддержания целостности мембраны (Chapin and Van Cleve, 1989). Во вторую ячейку добавляли 3 мкКи 32 P. Чтобы обнаружить возможные различия в транслокации между зрелыми и незрелыми зонами, два разных участка базального корня подвергали воздействию меченого фосфора клетки, содержащей радиоактивный источник.У одной группы растений обнаженный срез располагался на расстоянии 80 мм от кончика корня (рис. 2I). Во второй группе первый миллиметр от кончика корня базальных корней контактировал с меченой клеткой (рис. 2II). Через 4, 30 и 60 минут после добавления 32 P корни вырезали на нескольких участках. Базальные корни контактировали с раствором в каждой ячейке, и для выделения каждой ячейки использовали силиконовую смазку (Dow Corning, Мидленд, Мичиган, США). Во время воздействия меченого источника вся корневая система оставалась прикрепленной к побегу, а боковые корни не удалялись.Оставшаяся корневая система поддерживалась в растворе, эквивалентном тому, который использовался для заполнения ячеек из оргстекла. «Зона воздействия» (рис. 2) включала часть («ячейка с 32 P» на рис. 2I и II, длина 0,5 мм) корня, фактически погруженного в ячейку с 32 P и прилегающими 20 миллиметровые сегменты. «Зона измерения» включала в себя корневую часть 60 мм рядом с «зоной воздействия». После разделения корня на участки длиной 20 мм их ополаскивали в течение 4 мин в 1 мМ KH 2 PO 4 и 0.5 мМ CaSO 4 при 4 ° C для обмена 32 P из свободного пространства корня с последующей второй 4-минутной промывкой в 0,5 мМ CaSO 4 при 4 ° C (Chapin and Van Cleve, 1989). Радиоактивный фосфор в корневых срезах анализировали с помощью сцинтилляционного счетчика (Packard 1500, Packard Instruments Co, Downer’s Grove, IL, USA).
Рис. 2.
Схема метода отбора проб, используемого для оценки движения фосфора вдоль корневой оси.Два разных участка базального корня подвергались воздействию 32 P, расположенных на расстоянии 80 мм (I) или 10 мм (II) от кончика корня. Через 4, 30 и 60 мин после воздействия 32 P корни срезали на нескольких участках. «Зона воздействия» включала часть корня, погруженную в ячейку с радиоактивным источником (длина 0,5 мм) и прилегающие 20-миллиметровые сегменты). «Зона измерения» включала 60-миллиметровую корневую часть рядом с «зоной воздействия» и была вырезана с интервалом 2 мм для проведения измерений.
Рис. 2.
Схема метода отбора проб, использованного для оценки движения фосфора вдоль корневой оси. Два разных участка базального корня подвергались воздействию 32 P, расположенных на расстоянии 80 мм (I) или 10 мм (II) от кончика корня. Через 4, 30 и 60 мин после воздействия 32 P корни срезали на нескольких участках. «Зона воздействия» включала часть корня, погруженную в ячейку с радиоактивным источником (длина 0,5 мм) и прилегающие 20-миллиметровые сегменты).«Зона измерения» включала 60-миллиметровую корневую часть рядом с «зоной воздействия» и была вырезана с интервалом 2 мм для проведения измерений.
Приток фосфора
Чтобы оценить изменение притока фосфора вдоль оси различных классов корней фасоли обыкновенной, была разработана новая экспериментальная установка с количественной авторадиографией (рис. 3). Через 18 дней роста корневые системы осторожно вымывали из почвы, но оставляли прикрепленными к побегам и переносили в аэрированный раствор, содержащий 0.5 мМ CaSO 4. Корни оставались в этом растворе в течение 30–45 мин, а затем были помещены на 4 мин в пластиковые контейнеры, заполненные растворами для поглощения (100 мл) 0,5 мМ CaSO 4 и 1, 5 или 10 мкМ. KH 2 PO 4 с маркировкой 32 P (1 мкКи на контейнер). Для каждого раствора для поглощения отбирали 1 мл образца и анализировали на 32 P для расчета удельной активности. Средняя удельная активность составляла 1,4 мКи ммоль -1 P. Продолжительность периода воздействия на корни радиоактивного питательного вещества определяли после проведения эксперимента по транспортировке, описанного выше.После воздействия меченого раствора корневые системы ополаскивали в течение 4 минут в 1 мМ KH 2 PO 4 и 0,5 мМ CaSO 4 при 4 ° C для обмена 32 P из свободного пространства корня, после чего второй промывкой в течение 4 минут в 0,5 мМ CaSO 4 при 4 ° C. Затем корни отделяли от побега и помещали в сухой лед на 2 часа. Затем замороженные корневые системы помещали на ночь в лиофилизатор.
Рис. 3.
Схема эксперимента по притоку фосфора.
Рис. 3.
Схема эксперимента по притоку фосфора.
Оценка
32 Поглощение P по корневым осямДля каждой лиофилизированной корневой системы базальные корни, базальные боковые части и боковые части стержневого корня были осторожно иссечены у их основания и затем помещены на синюю бумагу для проращивания, выдерживая минимальное расстояние 2 см между ними, чтобы избежать перекрытия испускаемого излучения на пластине для визуализации. (Рис. 3). Эти корни сканировались с разрешением 1200 dpi с помощью WinRhizo v.4.0 (Regent Systems, Квебек, Канада) для измерения площади и длины поверхности с последующим экспонированием в течение ночи на фотостимулируемом люминофорном экране (Amersham Biosciences Corporation, Пискатауэй, Нью-Джерси, США). Экран фиксирует формирование скрытого изображения, вызванного ионизирующим излучением, исходящим от меченых корней. Затем экран генерирует фосфоресценцию (синий свет) пропорционально количеству радиоактивности в образце. В конце периода экспонирования экраны сканировали лазером с пространственным разрешением 100 мкм на пиксель и 16-битным диапазоном уровней серого с помощью системы PhosphorImager 445 SI (Amersham Biosciences Corporation, Пискатауэй, Нью-Джерси, США).Во время сканирования излучаемая экраном фосфоресценция собирается оптоволоконным жгутом и передается в фотоумножитель, и, наконец, получается графическое представление локализации поглощенных меченых ионов. Экспозиции в течение 10 часов было достаточно для получения точных изображений 32 P, накопленных мечеными корнями (рис. 4). Для количественной оценки накопления 32 P интересующий корневой сегмент был выбран путем рисования полосы на его изображении и проанализирован с помощью ImageQuant 5.0 (Amersham Biosciences Corporation, Пискатауэй, Нью-Джерси, США). Программа вычисляет площадь под кривой, созданной графиком зависимости интенсивности пикселей от местоположения пикселей вдоль полосы, и выражает ее в виде количества. Ширина полосы была выбрана оператором, а фоновая радиоактивность автоматически дисконтируется программой. При каждой экспозиции использовалась калибровочная кривая для преобразования данных, собранных из ImageQuant (выраженных в счетах), в мкКи. Градуировочная кривая построена для разных объемов раствора с известной радиоактивностью 32 P.Для каждой отдельной группы просканированных корней были получены таблицы подсчетов в зависимости от длины (расстояния) корней. Чтобы проверить точность беспленочной авторадиографии для обнаружения радиоактивности, результаты калибровочной кривой сравнивали с результатами аналогичной стандартной кривой, но были измерены стандартным методом для количественной оценки выбросов (сцинтилляционный счет).
Рис. 4.
Меченый фосфор абсорбируется отдельными сегментами корней бобов после 4 мин воздействия 1 мкМ фосфата, меченного 1 мкКи 32 P и подвергнутого воздействию люминесцентного экрана в течение 10 часов.Интенсивность серого пропорциональна накоплению фосфора. Изображение сканировали лазером с помощью беспленочной авторадиографической системы. Показана стандартная кривая для расчета удельной активности (вверху слева).
Рис. 4.
Меченый фосфор, абсорбированный отдельными сегментами корней фасоли после 4 минут воздействия 1 мкМ фосфата, меченного 1 мкКи 32 P и воздействия люминесцентного экрана в течение 10 часов. Интенсивность серого цвета пропорциональна накоплению фосфора.Изображение сканировали лазером с помощью беспленочной авторадиографической системы. Показана стандартная кривая для расчета удельной активности (вверху слева).
В примерах, приведенных в этой статье, корни были разделены на 2-миллиметровые сегменты, начиная от кончика корня, чтобы получить скорость притока фосфора. Пример расчета для отдельного сегмента прикорневых корней приведен в Приложении. Результаты также могут быть выражены по площади. В этом последнем случае требуются данные о площади поверхности каждого сегмента корня.Из ImageQuant было получено количество отсчетов через каждые 2 мм, а из результатов WinRhizo была измерена площадь поверхности корня в каждом сегменте 2 мм. Чтобы проверить, являются ли сегменты 2 мм эквивалентными в двух процедурах, сравнивали длину корневых сегментов, измеренную от кончика, полученного Imagequant и WinRhizo.
Чтобы получить точную количественную оценку и локализацию радионуклидов в корне, было проведено несколько предварительных экспериментов для определения соответствующей комбинации радиоактивности, продолжительности периода контакта между корнями и меченым раствором и продолжительности периода воздействия корень к люминофорному экрану.Если накопление 32 P в корне не находится в оптимальном диапазоне обнаружения, либо недоэкспонирование, либо перенасыщение изображения будут проблематичными. Основываясь на этих предварительных экспериментах, условия, использованные в основном эксперименте: 1 мкКи в 100 мл 1–10 мкМ KH 2 PO 4 , 4-минутный контакт с меченым источником и 10-часовое воздействие на корень люминофорный экран. Если вся корневая система подвергается воздействию люминесцентного экрана, скорость поглощения отдельных сегментов корней невозможно рассчитать из-за перекрытия излучения, испускаемого соседними сегментами корня.Это причина того, что отдельные сегменты корня отделялись друг от друга после контакта с меченым фосфором и перед воздействием на люминесцентный экран. Диапазон интенсивности серого на рис. 4 представляет различные значения притока.
Статистический анализ
В эксперименте по притоку 30 растений (две обработки фосфором в течение периода роста тремя концентрациями фосфора в питательном растворе на пять блоков) были расположены по полностью рандомизированной схеме блоков.Для каждого растения усредняли результаты по нескольким (2–8) единицам каждого корневого класса. ANOVA с повторными измерениями и регрессионный анализ выполняли с использованием SYSTAT v. 8.0 (SPSS Inc., Эванстон, Иллинойс, США).
Результаты
Точность беспленочной авторадиографии
Тесная взаимосвязь ( r 2 = 0,99) (рис. 5) между количеством отсчетов стандартной кривой (как показано на рис. 4), измеренной программой ImageQuant, и количеством радиоактивности, измеренной сцинтилляционным счетом, показывает что цифровая авторадиография может дать точное значение радиоактивности 32 P, присутствующей в образце.На «пространственную» точность беспленочной авторадиографии для количественной оценки притока фосфора с высоким пространственным разрешением (каждые 2 мм длины высушенного корня) указывает почти точное соотношение ( r 2 = 0,99) (рис. длина сегментов корня, полученных из изображений радиоактивности (как на рис. 4), и длина того же корня, измеренная с помощью WinRhizo.
Рис. 5.
Соотношение между количеством отсчетов стандартной кривой, как показано на Рис.3 измерено цифровой авторадиографией и стандартным методом (жидкостный сцинтилляционный счет).
Рис. 5.
Зависимость между количеством отсчетов стандартной кривой, как показано на рис. 3, измеренных с помощью цифровой авторадиографии и стандартным методом (жидкостный сцинтилляционный счет).
Рис. 6.
Взаимосвязь между длиной корня, измеренной путем анализа цифровых авторадиографических изображений ионизирующего излучения, испускаемого корнями (как на Рис.3) и длину корня, проанализированную программным обеспечением для анализа корней (WinRhizo).
Рис. 6.
Взаимосвязь между длиной корня, измеренной путем анализа цифровых авторадиографических изображений ионизирующего излучения, испускаемого корнями (как на рис. 3), и длиной корня, проанализированной с помощью программного обеспечения для анализа корней (WinRhizo) .
Осевой перенос фосфора : После 4 мин воздействия на корни указанного источника фосфора доля фосфора, переместившаяся в «зону измерения» (рис.2) представляли 0,05–0,12% фосфора, обнаруженного в подвергнутом воздействию сегменте (Таблица 1). Когда период воздействия был увеличен до 60 минут, доля переносимого фосфора увеличилась почти до 25% (таблица 1). В каждом случае базипетальное движение было больше, чем акропетальное движение. Был сделан вывод, что меченый фосфор, обнаруженный в данном участке корня после 4 минут воздействия, точно отражает количество притока фосфора в этот участок корня. Таким образом, в эксперименте с чистым притоком фосфора корни подвергались воздействию меченого источника фосфора в течение 4 мин.
Таблица 1.Влияние продолжительности воздействия и положения маркировки на базипетальное (то есть по направлению к основанию корня) и акропетальное (то есть по направлению к кончику корня) движение 32 P
Время выдержки до 32 P (мин) . | Положение надписи (мм от кончика корня) . | Поглощение P в отмеченной области (пмоль P г −1 сырой массы мин. −1 ) . | Базипетальное движение P (% P, обнаруженного в помеченной области) . | Акропетальное движение фосфора (% фосфора, обнаруженного в помеченной области) . | ||
|---|---|---|---|---|---|---|
| 4 | 80 | 100 (23) | 0,05 (0,02) | ND a | ||
| 30 | 80 | 27 (4,3) | 0,05 (0,01) | |||
| 4 | 10 | 99 (25) | 0.12 (0,05) | NA b | ||
| 30 | 10 | 43 (19) | 1,91 (0,55) | NA | ||
| 60 | 80 55 | 24,17 (7,21) | 0,10 (0,02) |
Время выдержки до 32 P (мин) . | Положение надписи (мм от кончика корня) . | Поглощение P в отмеченной области (пмоль P г −1 сырой массы мин. −1 ) . | Базипетальное движение P (% P, обнаруженного в помеченной области) . | Акропетальное движение фосфора (% фосфора, обнаруженного в помеченной области) . | |||||
|---|---|---|---|---|---|---|---|---|---|
| 4 | 80 | 100 (23) | 0,05 (0,02) | ND a | |||||
| 30 | 80 | 27 (4.3) | 0,37 (0,22) | 0,05 (0,01) | |||||
| 4 | 10 | 99 (25) | 0,12 (0,05) | NA b | |||||
| 30 43 (19) | 1,91 (0,55) | NA | |||||||
| 60 | 80 | 55 (21) | 24,17 (7,21) | 0,10 (0,02) | |||||
Время воздействия 32 P (мин) . | Положение надписи (мм от кончика корня) . | Поглощение P в отмеченной области (пмоль P г −1 сырой массы мин. −1 ) . | Базипетальное движение P (% P, обнаруженного в помеченной области) . | Акропетальное движение фосфора (% фосфора, обнаруженного в помеченной области) . | ||
|---|---|---|---|---|---|---|
| 4 | 80 | 100 (23) | 0,05 (0,02) | ND a | ||
| 30 | 80 | 27 (4,3) | 0,05 (0,01) | |||
| 4 | 10 | 99 (25) | 0,12 (0,05) | NA b | ||
| 30 | 10 | 43 (19) 1 | 91 (0,55)NA | |||
| 60 | 80 | 55 (21) | 24,17 (7,21) | 0,10 (0,02) |
Положение надписи (мм от кончика корня) . | Поглощение P в отмеченной области (пмоль P г −1 сырой массы мин. −1 ) . | Базипетальное движение P (% P, обнаруженного в помеченной области) . | Акропетальное движение фосфора (% фосфора, обнаруженного в помеченной области) . | |||
|---|---|---|---|---|---|---|
| 4 | 80 | 100 (23) | 0,05 (0,02) | ND a | ||
| 30 | 80 | 27 (4,3) | 0,05 (0,01) | |||
| 4 | 10 | 99 (25) | 0.12 (0,05) | NA b | ||
| 30 | 10 | 43 (19) | 1,91 (0,55) | NA | ||
| 60 | 80 55 | 24,17 (7,21) | 0,10 (0,02) |
Приток фосфора
Базальные боковые стороны и боковые части стержневого корня были около 20 мм в длину, а базальные корни были примерно в десять раз длиннее.Для прямого сравнения трех классов корней на рис. 7 показан общий приток фосфора в верхушечные 20 мм каждого класса корней. Среди классов корней наибольший приток наблюдался у базальных: они поглощали в 2,5 и 1,9 раза больше фосфора, чем классы боковых и базальных боковых корней соответственно (рис. 7; табл. 2). Более высокая очевидная эффективность базальных корней наблюдалась у обоих растений, которые были выращены в условиях с высоким и низким содержанием фосфора и при трех концентрациях фосфора, предлагаемых в меченых питательных растворах (рис.7). У базальных боковых и центральных корневых каналов поступление фосфора было сопоставимым (рис. 7). Единственные различия между этими двумя классами были обнаружены у растений, выращенных в условиях низкого содержания фосфора и подвергшихся воздействию двух самых низких концентраций в меченых растворах (1 мкМ и 5 мкМ). В этих обработках базальные отводы показали более высокий приток общего фосфора, чем отводы от стержневых корней (рис. 7). Концентрация фосфора в меченом питательном растворе была фактором, оказавшим наибольшее влияние на приток фосфора к растениям (рис.7; Таблица 2). В среднем приток фосфора был в 6,9 раза и в 2,1 раза больше при максимальной концентрации (10 мкМ), чем при 1 мкМ и 5 мкМ соответственно.
Рис. 7.
Скорость поступления фосфора в первые 20 мм (от кончика корня) базального (закрытые столбцы), бокового базального (открытые столбцы) и бокового ответвления (столбики в стопке) корней фасоли обыкновенной. Растения выращивали в течение 18 дней при низком (1 мкМ) или высоком (30 мкМ) количестве фосфора, а затем подвергали в течение 4 мин воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P.Значения представляют собой среднее значение для пяти растений и выражены на основе сухого корня. Для каждого растения усредняли результаты по нескольким (2–8) единицам каждого корневого класса. Столбцы представляют собой SE средних значений. Цифры над столбиками указывают соотношение притока между высоким и низким статусом фосфора.
Рис. 7.
Скорость поступления фосфора в первые 20 мм (от кончика корня) базального (закрытые столбцы), боковых базальных (открытые столбцы) и боковых ответвленных (столбики) корней фасоли обыкновенной.Растения выращивали в течение 18 дней при низком (1 мкМ) или высоком (30 мкМ) количестве фосфора, а затем подвергали в течение 4 мин воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P. Значения представляют собой среднее из пяти значений. растения и выражаются на основе сухих корней. Для каждого растения усредняли результаты по нескольким (2–8) единицам каждого корневого класса. Столбцы представляют собой SE средних значений. Цифры над столбиками указывают соотношение притока между высоким и низким статусом фосфора.
Таблица 2.Таблица дисперсионного анализа эксперимента по чистому притоку фосфора с фасолью обыкновенной
. | степеней свободы . | F Значение и значение a . | . | . | |||||
|---|---|---|---|---|---|---|---|---|---|
. | . | Приток фосфора . | Приток фосфора (по площади) . | Приток фосфора (по длине) . | |||||
| Тип корня (A) | 2 | 9,05 *** | 12,88 *** | 87,06 *** | |||||
| Phosphor6 | 0,7 NS | 10,83 ** | 7,22 ** | ||||||
| Положение (C) | 9 | NA | 0.33 NS | 2,85 ** | |||||
| Pconc (D) | 2 | 20,83 *** | 256,83 *** | 197,32 *** | 2 | 0,11 NS | 0,80 NS | 1,25 NS | |
| A × C | 18 | NA | 0,59 NS | 0,53 NS | |||||
| 4.21 ** | 13,80 *** | ||||||||
| B × C | 9 | NA | 0,06 NS | 0,07 NS | |||||
| B × D | 2 | 3.20 *7.80 *** | |||||||
| C × D | 18 | NA | 0.58 NS | 1.03 NS | 903степеней свободы . | F Значение и значение a . | . | . | |
. | . | Приток фосфора . | Приток фосфора (по площади) . | Приток фосфора (по длине) . | |||||
| Тип корня (A) | 2 | 9,05 *** | 12,88 *** | 87,06 *** | |||||
| Phosphor6 | 0,7 NS | 10,83 ** | 7,22 ** | ||||||
| Положение (C) | 9 | NA | 0,33 NS | 2,85 ** | 2 | 20.83 *** | 256,83 *** | 197,32 *** | |
| A × B | 2 | 0,11 NS | 0,80 NS | 1,25 NS | 18 | NA | 0,59 NS | 0,53 NS | |
| A × D | 4 | 1,43 NS | 4,21 ** | 13,804 *** | 13,804 *** | К9 | NA | 0.06 NS | 0,07 NS |
| B × D | 2 | 0,85 NS | 3,20 * | 7,80 *** | |||||
| C × D | 0,58 NS | 1,03 NS | |||||||
Таблица ANOVA эксперимента по чистому притоку фосфора с растениями фасоли обыкновенной
. | степеней свободы . | F Значение и значение a . | . | . | |||||
|---|---|---|---|---|---|---|---|---|---|
. | . | Приток фосфора . | Приток фосфора (по площади) . | Приток фосфора (по длине) . | |||||
| Тип корня (A) | 2 | 9,05 *** | 12,88 *** | 87,06 *** | |||||
| Phosphor6 | 0,7 NS | 10,83 ** | 7,22 ** | ||||||
| Положение (C) | 9 | NA | 0,33 NS | 2,85 ** | 2 | 20.83 *** | 256,83 *** | 197,32 *** | |
| A × B | 2 | 0,11 NS | 0,80 NS | 1,25 NS | 18 | NA | 0,59 NS | 0,53 NS | |
| A × D | 4 | 1,43 NS | 4,21 ** | 13,804 *** | 13,804 *** | К9 | NA | 0.06 NS | 0,07 NS |
| B × D | 2 | 0,85 NS | 3,20 * | 7,80 *** | |||||
| C × D | 0,58 NS | 1,03 NS | |||||||
. | степеней свободы . | F Значение и значение a . | . | . | ||||
|---|---|---|---|---|---|---|---|---|
. | . | Приток фосфора . | Приток фосфора (по площади) . | Приток фосфора (по длине) . | ||||
| Тип корня (A) | 2 | 9,05 *** | 12,88 *** | 87.06 *** | ||||
| Фосфор (B) | 1 | 0,7 NS | 10,83 ** | 7,22 ** | ||||
| NA | 9 Положение (C) | 9 | 0,33 NS | 2,85 ** | ||||
| Pconc (D) | 2 | 20,83 *** | 256,83 *** | 197.32 *** | × В2 | 0.11 NS | 0,80 NS | 1,25 NS |
| A × C | 18 | NA | 0,59 NS | 0,53 NS | ||||
| A × D | 4 | 1.45 * | 13,80 *** | |||||
| B × C | 9 | NA | 0,06 NS | 0,07 NS | ||||
| B × D | 2 | 0,8 | 7.80 *** | |||||
| C × D | 18 | NA | 0,58 NS | 1,03 NS | ||||
Дисперсионный анализ показал значительные эффекты корневого класса и растений на притоке фосфора, выраженном на основе длины корня (т. е. пмоль P мм -1 мин -1 ) или на основе площади корня (пмоль P см -2 мин -1 ) (Таблица 2). Наблюдались интерактивные эффекты между статусом фосфора и концентрацией фосфора в маркированном питательном растворе: рисунки 8 и 9 показывают, что фосфорный стресс влиял на приток только тогда, когда растения подвергались воздействию низких концентраций фосфора (см. Также таблицу 2).Влияние положения сегмента корня вдоль оси корня было статистически значимым, когда приток фосфора был выражен на основе длины корня, но не когда выражен на основе площади корня (Таблица 2). Когда приток фосфора выражался по длине, у базальных корней приток уменьшался в тех частях корня, которые расположены близко к кончику корня (рис. 8, 9). Этот эффект был статистически значимым для растений, выращенных как в условиях с низким, так и с высоким содержанием фосфора, подвергшихся воздействию 1 мкМ меченого фосфора (рис. 8, 9).Максимальный приток на единицу длины в базальных корнях наблюдался на участках, расположенных в 20 мм от кончика корня. При расчете притока фосфора вдоль корневой оси базальных корней на основе площади не наблюдалось последовательной пространственной картины (рис. 8, 9). Приток фосфора был довольно постоянным в сегментах базального корня рядом с гипокотилем (который представляет самые старые сегменты этого класса корней) (рис. 10) и не отличался от сегментов, расположенных в 20–80 мм от кончика корня, как показано на Рис 8 и 9.Для бокового главного корня и боковых базальных корней не наблюдалось явного влияния положения корневого среза вдоль оси корня, если оно выражено как площадь или основание длины (рис. 8, 9).
Рис. 8.
Скорость поступления фосфора по оси базального, бокового базального и бокового стержневых корней фасоли обыкновенной. Левая колонка: приток на единицу площади поверхности корня; правый столбец: поглощение на единицу длины корня. Растения выращивали в течение 18 дней при низком содержании фосфора (1 мкМ фосфата), а затем подвергали в течение 4 минут воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P.Значения представляют собой среднее значение для пяти растений и выражены на основе сухого корня. Для каждого растения усредняли результаты по нескольким (2–8) единицам каждого корневого класса. Столбцы представляют собой критическое значение ( P <0,05) для сравнения средних значений каждого корневого класса в соответствии с тестом LSD.
Рис. 8.
Скорость поступления фосфора по оси базального, бокового базального и бокового стержневых корней фасоли обыкновенной. Левая колонка: приток на единицу площади поверхности корня; правый столбец: поглощение на единицу длины корня.Растения выращивали в течение 18 дней при низком содержании фосфора (1 мкМ фосфата), а затем подвергали в течение 4 минут воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P. Значения представляют собой среднее значение для пяти растений и выражены на сухая корневая основа. Для каждого растения усредняли результаты по нескольким (2–8) единицам каждого корневого класса. Столбцы представляют собой критическое значение ( P <0,05) для сравнения средних значений каждого корневого класса в соответствии с тестом LSD.
Фиг.9.
Скорость поступления фосфора по оси базального, бокового базального и бокового стержневых корней фасоли обыкновенной. Левая колонка: приток на единицу площади поверхности корня; правый столбец: поглощение на единицу длины корня. Растения выращивали в течение 18 дней при высоком содержании фосфора (30 мкМ фосфата), а затем подвергали в течение 4 минут воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P. Значения представляют собой среднее значение для пяти растений и выражены на сухая корневая основа. Для каждого растения усредняли результаты по нескольким (2–8) единицам каждого корневого класса.Столбцы представляют собой критическое значение ( P <0,05) для сравнения средних значений каждого корневого класса в соответствии с тестом LSD.
Рис. 9.
Скорость поступления фосфора по оси базального, бокового базального и бокового стержневых корней фасоли обыкновенной. Левая колонка: приток на единицу площади поверхности корня; правый столбец: поглощение на единицу длины корня. Растения выращивали в течение 18 дней при высоком содержании фосфора (30 мкМ фосфата), а затем подвергали в течение 4 минут воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P.Значения представляют собой среднее значение для пяти растений и выражены на основе сухого корня. Для каждого растения усредняли результаты по нескольким (2–8) единицам каждого корневого класса. Столбцы представляют собой критическое значение ( P <0,05) для сравнения средних значений каждого корневого класса в соответствии с тестом LSD.
Рис. 10.
Скорость поступления фосфора по оси базальных корней фасоли обыкновенной, начиная от основания корня (где базальный корень соединяется с осью основного корня).Растения выращивали в течение 18 дней при низком (1 мкМ, левая колонка) или высоком (30 мкМ, правая колонка) подаче фосфора, а затем подвергали в течение 4 мин воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P. Значения представляют собой среднее значение для пяти растений и выражены на основе сухого корня. Для каждого растения усредняли результаты по нескольким (2–8) прикорневым корням. Столбцы представляют собой SE средних значений.
Рис. 10.
Скорость поступления фосфора по оси базальных корней фасоли обыкновенной, начиная с основания корня (где базальный корень соединяется с осью основного корня).Растения выращивали в течение 18 дней при низком (1 мкМ, левая колонка) или высоком (30 мкМ, правая колонка) подаче фосфора, а затем подвергали в течение 4 мин воздействию 1, 5 и 10 мкМ фосфата, меченного 1 мкКи 32 P. Значения представляют собой среднее значение для пяти растений и выражены на основе сухого корня. Для каждого растения усредняли результаты по нескольким (2–8) прикорневым корням. Столбцы представляют собой SE средних значений.
Обсуждение
Цифровая авторадиография
Цифровая авторадиография позволила составить пространственное картирование способности притока фосфора вдоль корневых осей растений фасоли обыкновенной.Не нужно отделять корни от побега, что является общим требованием многих современных аналитических методов (Chapin and Van Cleve, 1989). Удаление корня может повлиять на приток фосфора, потому что (i) потребность побегов является основной движущей силой поглощения питательных веществ, и этот сигнал не может быть обнаружен удаленными корнями; (ii) удаление корня приводит к быстрой потере углеводов, что снижает усвоение питательных веществ (Bloom and Caldwell, 1988). Для кальция Уайт и др. . (1992) обнаружили, что чистый приток неповрежденных корней был почти в 8 раз больше, чем приток удаленных корней.
Поскольку люминофорный экран эффективно обнаруживает радиоактивность в ограниченном диапазоне, при использовании цифровой авторадиографии следует соблюдать некоторые меры предосторожности. Если эксперименты проводятся с использованием специфической активности, которая не приводит к накоплению 32 P в оптимальном диапазоне обнаружения, либо недодержка, либо перенасыщение изображения не позволят правильно определить количество и локализацию присутствующих меченых ионов. в корнях. Чтобы избежать этой проблемы, необходимо тщательно отрегулировать уровень радиоактивности в питательном растворе и продолжительность воздействия на экран.Низкая радиоактивность (или очень короткое время воздействия) приводит к недостаточной экспозиции экрана, из-за чего трудно отличить фактический приток питательных веществ от естественного фонового излучения. Чрезмерная радиоактивность или очень длительные периоды воздействия приводят к большей экспериментальной ошибке из-за перенасыщения изображения. Перенасыщение приводит к появлению темных пятен на люминофорном экране, по которым невозможно оценить реальные размеры анализируемого участка корня. Перед использованием этого метода следует провести предварительные эксперименты, чтобы определить наиболее удобную комбинацию (i) количества радиоактивности меченого раствора, (ii) продолжительности периода контакта корня с меченым раствором и ( iii) продолжительность периода воздействия на меченый корень люминофорным экраном.В этом случае использовали 1 мкКи в 100 мл 1–10 мкМ KH 2 PO 4 в течение 4 мин и 10 ч соответственно. Как правило, время воздействия меченого раствора должно быть достаточно продолжительным, чтобы питательные вещества прошли через свободное пространство апоплазмы и достигли симплазмы, и должно быть достаточно коротким, чтобы избежать переноса питательного вещества в соседние корневой сегмент. Чтобы определить продолжительность периода поглощения, необходимо принять во внимание длину самого маленького сегмента корня, который необходимо проанализировать.Например, если за единицу анализа взять 2 мм корня, продолжительность периода абсорбции должна быть достаточно короткой, чтобы избежать перемещения 32 P на расстояние 2 мм. Для определения скорости перемещения можно провести простые эксперименты, такие как метод смежных камер, описанный в этой статье. Радиоактивность питательного раствора и время воздействия на люминофорный экран следует регулировать одновременно. Описанный здесь метод может быть полезен для других радионуклидов, важных для биологии растений, включая 33 P, 45 Ca, 35 S, 14 C, 3 H и 36 Cl.Параметры метода (в основном радиоактивность и время воздействия) должны быть адаптированы к энергетическим характеристикам каждого радионуклида.
Поступление фосфора по оси различных классов корней фасоли обыкновенной
В процессе поглощения питательных веществ необходимо различать четыре различных процесса: приток, накопление, перемещение и отток. Последний процесс считается актуальным только в условиях низкой доступности питательных веществ (Elliot et al ., 1984; Джексон и др. ., 1976). Приток связан с активностью носителей, для которой характерна классическая ферментативная кинетика (Epstein, 1972; см. Также Nissen, 1996). Хранение связано с наличием клеточных вакуолей и, следовательно, с участками корня с некоторой степенью созревания (Lee and Ratcliffe, 1993). Транслокация также требует определенного уровня созревания, поскольку она зависит от зрелых элементов ксилемы и флоэмы. В этой статье для количественной оценки притока независимо от транслокации использовался 4-минутный период воздействия меченого раствора, в течение которого происходила незначительная транслокация.Когда время экспозиции было увеличено до 60 мин, большое количество фосфора было доставлено базипетально к побегу из участков корня, расположенных в 80 мм от кончика корня, где ксилема полностью созрела. Эти результаты, как и предыдущие отчеты, сосредоточены на движении недавно поглощенных ионов. Было бы интересно исследовать, есть ли какие-либо различия в перемещении недавно и ранее поглощенных ионов.
Было замечено, что дефицит фосфора увеличивает способность поглощения фосфора, что согласуется с хорошо известным фактом, что нутритивный статус оказывает важное влияние на поглощение фосфора (Aerts and Chapin, 2000; Cartwright, 1972; Clarkson and Scattergood, 1982; Clarkson и др. ., 1978; Дрю и др. ., 1984). Эта модуляция составляет эффективную регуляцию обратной связи и сильно различается у разных видов (Cogliatti and Clarkson, 1983; Drew et al ., 1984; Jungk et al ., 1990). В этом исследовании было замечено, что это регулирование не было согласованным ни среди классов корней, ни среди концентраций фосфора в питательном растворе. Дефицит фосфора увеличивал приток фосфора только в базальные корни и их боковые стороны. Есть несколько предшественников такого рода о различном влиянии статуса питания на различные классы корней растений, выращиваемых в однородных средах (Russel and Sanderson, 1967; Waisel and Eshel, 1992).Напротив, в гетерогенных средах с ионами фосфора, сконцентрированными в участках, увеличение способности поглощать фосфор корнями, расположенными на участке, было хорошо задокументировано (Jackson et al ., 1990; Snapp et al ., 1995). . Что касается интерактивных эффектов состояния питания растений и концентраций ионов в питательном растворе, сообщалось о двух различных паттернах реакции (Картрайт, 1972; Дрю и др. ., 1984; Данлоп и др. ., 1997). Различные ответы были связаны с изменениями в кинетике механизма захвата ионов: увеличение В макс (т.е. максимальная скорость поглощения) без заметных изменений К м (т.е. концентрация ионов субстрата, дающая половину В. max ) были признаны ответственными за повышенный приток фосфора, вызванный фосфорным стрессом у ячменя (Drew et al ., 1984) и Arabidopsis (Dunlop et al ., 1997). В отличие от этого Картрайт (1972) обнаружил, что V max оставалось относительно постоянным, но K m уменьшалось в ячмене с недостатком фосфора. В этом последнем отчете различия в притоке фосфора между стрессируемыми фосфором и фосфоро-достаточными растениями были меньше при более высоких концентрациях фосфора в питательном растворе из-за отсутствия корректировок в V max . Эти результаты предполагают, что влияние питательного статуса растения на приток фосфора снижается при более высоких концентрациях питательных веществ в почвенном растворе.Результаты этого исследования явно более совместимы с этой последней моделью, чем с первой, поскольку различия между контрольными и подвергнутыми стрессу растениями обнаруживались при концентрации фосфора 1 мкМ и 5 мкМ, но не при 10 мкМ фосфора (рис. 7).
Базальные корни характеризовались большей интенсивностью поступления фосфора, оцениваемой по длине, чем боковые корни. Это наблюдалось во всех использованных экспериментальных режимах. Когда приток измерялся на основе площади, различия между классами корней были меньше, хотя базальные корни оставались группой с самым высоким притоком фосфора при низких концентрациях фосфора в поглощающем растворе.Самая низкая концентрация фосфора, использованная в этом исследовании (1 мкМ), соответствует типичной концентрации фосфора в почвенном растворе в большинстве полевых условий (Barber, 1995). Большее поглощение фосфора базальными корнями может быть полезным признаком для генотипов, у которых базальные корни и их боковые части сосредоточены в верхнем слое почвы, например BAT 477, генотип, использованный в эксперименте этого исследования (Bonser et al ., 1996; Liao ). et al ., 2001). Было замечено, что у этого генотипа почти 40% базальных корней (и их боковых сторон) сосредоточено в верхнем слое почвы (Liao et al ., 2001). Поскольку доступность фосфора обычно наиболее высока в верхнем слое почвы и уменьшается с глубиной (Chu and Chang, 1966; Pothuluri et al ., 1986), генотипы с неглубокими базальными корнями обладают большей способностью усваивать фосфор, что наблюдается в теплице и в поле (Линч и Браун, 2001; Рубио и др. ., 2003). Хотя основное внимание в настоящей работе уделяется отдельным корням, грубая оценка способности притока фосфора для всего растения может быть сделана с использованием данных о притоке фосфора, полученных здесь, и данных о длине различных классов корней у 14-дневных растений фасоли. (резюме.Carioca, общая длина корней 35,14 м), о которых сообщалось ранее (Lynch and van Beem, 1993). Пропускная способность 3,8 нмоль P min -1 растение -1 оценивается с использованием обоих источников информации, если в качестве примера рассматриваются корни, контактирующие с 5 мкМ раствором P. Скорости поглощения при данной внешней концентрации питательных веществ могут быть получены из опубликованных значений кинетики поглощения ( K m , V max , C min ). Например, из данных, рассмотренных Tinker and Nye (2000; см. Их Таблицу 5.4), скорости поглощения 5,5, 2,8 и 6,1 нмоль P min -1 растение -1 можно оценить для лука, рапса и ячменя соответственно. Для Paspalum dilatatum , дикого растения, коэффициент 1,7 нмоль P min -1 растение -1 можно оценить по данным, опубликованным Rubio et al . (1997). Для расчета этих показателей общая длина корня и внешняя концентрация фосфора были приведены к значениям, используемым для примера с обычными бобами (35,14 м и 5 мкМ P).
Было высказано предположение, что фосфор либо достаточно равномерно абсорбируется по всей поверхности корней однолетних растений (Bowen, 1969; Rovira, Bowen, 1970; Russel and Clarkson, 1975; Russel and Sanderson, 1967), либо наиболее активно всасывается в апикальные области корневой оси (Clarkson et al ., 1978). В соответствии с первой группой отчетов, относительно однородные скорости притока наблюдались вдоль корневых осей. Однако базальные корни показали неоднородный характер притока фосфора по осям корня, но только тогда, когда приток рассчитывался на основе длины корня. Тот факт, что эта пространственная неоднородность исчезла, когда приток был рассчитан на основе площади, указывает на то, что диаметр корня является основным фактором, регулирующим характер притока вдоль корневой оси. Эти результаты нельзя напрямую сравнивать с большинством опубликованных отчетов, поскольку в подавляющем большинстве из них учитываются более длительные периоды воздействия указанного источника, обычно более 20 минут, а в некоторых случаях до 24 часов (Clarkson et al ., 1978). Как отмечалось выше, в такой период времени не только приток, но и процессы транслокации ответственны за характер локализации фосфора в различных частях корня. Эксперименты с длительным воздействием являются надежными индикаторами всех процессов, связанных с усвоением питательных веществ, действующих совместно, и способности различных частей корня экспортировать фосфор от корня к побегу. Однако они не подходят для определения пространственной картины притока фосфора.В результатах этого исследования вариации диаметра корня по оси корня объясняли различия в притоке на единицу длины, поскольку не было обнаружено больших различий, когда результаты выражались на единицу площади поверхности. Подобную тенденцию наблюдали Рассел и Сандерсон (1967), которые проанализировали локализацию фосфора в корневой оси после 24-часового воздействия меченого источника. Существенная однородность в структуре притока фосфора, обнаруженная вдоль корневой оси, выраженная таким образом, может быть связана с тем фактом, что все растения в этом исследовании были молодыми (18 дней) и все их корни активно росли.Однако интересно отметить, что не было обнаружено существенных различий в притоке между окрестностями вершины и основанием базальных корней. Учитывая, что базальные корни выходят из гипокотиля через 3-4 дня после прорастания, разница в возрасте между кончиком и основанием базальных корней растений составляла примерно 2 недели. В отличие от кукурузы, Gao et al . (1998) наблюдали резкое снижение способности поглощать фосфор между 5-м и 10-м днем роста корней, в то время как более старые корни демонстрировали более постепенное снижение.Результаты этого исследования согласуются с недавними открытиями, показывающими, что переносчики фосфора присутствуют во всех частях корневой системы, несмотря на анатомические различия и уровни созревания различных частей (Muchhal and Raghothama, 1999). В поле фактическая картина притока фосфора в корневую систему будет результатом взаимодействия способности притока фосфора с пространственной структурой доступности фосфора в почве. В этом контексте рисунки 7, 8 и 9 иллюстрируют большую зависимость притока фосфора от концентрации фосфора в почвенном растворе.Можно ожидать, что новейшие сегменты корневой системы (близкие к верхушке) в поле будут сталкиваться с почвенными доменами, более богатыми фосфором, чем самые старые сегменты, в которых ризосфера уже обеднена фосфором.
Выводы
Метод, описанный в этой статье, предлагает удобный протокол для изучения поступления фосфора в корни. Метод использует цифровую авторадиографию для визуализации и количественной оценки притока фосфора в неповрежденные корневые системы.Это позволяет точно определить пространственный характер притока фосфора. Следуя протоколу, описанному здесь, карты чистой притока питательных веществ и кинетических параметров питательных веществ могут быть получены и применены для изучения широкого круга вопросов. Эта информация может помочь в построении моделей поглощения питательных веществ, указав, что для молодых растений разумно предположить постоянный приток фосфора вдоль корневой системы, но необходимо учитывать различия между классами корней.
Приложение
Пример расчета притока фосфора для отдельного сегмента корня
Индивидуальный: завод № 56
Тип корня: основной корень
Анализируемый сегмент корня: 0–2 мм (от кончика корня)
Обработка фосфором: высокая
Концентрация поглощающего раствора: 1 мкМ P
Радиоактивность поглощающего раствора: 7573 имп / мин мл -1 (измерено сцинтилляционным счетом).
Объем поглощаемого раствора: 180 мл
Общая радиоактивность: 7573 × 180 = 1363 140 имп / мин в 180 мл
Завод №56 был отнесен к 1 мкМ P, следовательно, в объеме 180 мл содержится 0,18 мкмоль P
Удельная активность равна импульсов в минуту 32 P мкмоль -1 P: 1363 140 / 0,18 = 7 573 000 импульсов в минуту мкмоль −1 P
Измерение с помощью Pimager базального корня № 56 при 0–2 мм: 441,2 отсчета
Преобразование отсчетов Pimager в число импульсов в минуту путем калибровки стандартной кривой (эта калибровка выполняется для каждой группы корней, как показано на рис. 4):
Для растений № 56: импульсов в минуту = 0,023 × отсчетов Pimager –0.29;
, так как counts = 441,2, то имп / мин = 9,857
Превращение радиоактивности (имп / мин) в мкмоль фосфора по удельной активности:
9,857 имп / мин / 7 573 000 имп / мин мкмоль -1 P = 1,29 × 10 -6 мкмоль P
мкмоль P в первых 2 мм: 1,29 × 10 −6 мкмоль P
Время воздействия питательного раствора: 4 мин
Приток P в минуту = 1,29 × 10 −6 /4
P скорость притока: 3,23 × 10 −7 мкмоль P мм −1 мин −1
Скорость притока P: 0.323 пмоль P мм −1 мин −1
Мы признательны за поддержку со стороны гранта USDA / NRI 92 и гранта NSF IBN-0135872 JPL и KM Brown. Мы благодарим Роберта Снайдера за его техническую помощь и Роберта Т. Симпсона за разрешение доступа к его лабораторным помещениям.
Список литературы
Aerts R, Chapin III FS.
2000
. Еще раз о минеральном питании дикорастущих растений: переоценка процессов и закономерностей.Достижения в области экологических исследований
30
,1
–67.Aguirrezabal LAN, Pellerin S, Tardieu F.
1993
. Углеродное питание, ветвление и удлинение корней: может ли нынешний уровень знаний позволить использовать прогностический подход на уровне всего растения?Экологическая и экспериментальная ботаника
33
,121
–130.Барбер С.
1995
. Биодоступность питательных веществ в почве: механистический подход. Нью-Йорк: Дж. Вили и сыновья.Bar-Yosef B, Kafkaki U, Bresler E.
1972
. Поглощение фосфора растениями в полевых условиях. I. Теоретическая модель и экспериментальное определение параметров.Почвоведение
36
,783
–800.Блум А., Колдуэлл, РМ.
1988
. Удаление корней снижает всасывание питательных веществ и потоки газов.Физиология растений
87
,794
–796.Bonser AM, Lynch JP, Snapp S.
1996
. Влияние дефицита фосфора на угол роста базальных корней у Phaseolus vulgaris .Новый фитолог
132
,281
–288.Bowen GD.
1969
. Поглощение ортофосфата и его включение в органические фосфаты корнями Pinus radiata .Австралийский журнал биологических наук
22
,1125
–1135.Берли Дж., Нвоке ФИО, Лейстер Дж., Попхэм Р.
1970
. Связь созревания ксилемы с абсорбцией и перемещением P.Американский журнал ботаники
57
,504
–511.Картрайт Б.
1972
. Влияние дефицита фосфата на кинетику поглощения фосфата стерильными иссеченными корнями ячменя и некоторые факторы, влияющие на дефицит поглощения ионов.Почвоведение и анализ растений
3
,313
–322.Чапин III ФС, Ван Клив К.
1989
. Подходы к изучению поглощения, использования и потери питательных веществ растениями. В: Pearcy RW, Ehleringer J, Mooney HA, Rundel PW. Физиологическая экология растений: полевые методы и приборы . Лондон: Чепмен и Холл, 185–207.Чу В.К., Чанг СК.
1966
. Поверхностная активность неорганического почвенного фосфора.Почвоведение
101
,459
–464.Clarkson DT.
1996
. Структура корня и места захвата ионов. В: Waisel Y, Eshel A, Kafkafi U, eds. Корни растений, скрытая половина. Нью-Йорк: Марсель Деккер Инк, 483–510.Clarkson DT, Sanderson J, Scattergood CB.
1978
. Влияние фосфатного стресса и поглощения и транслокации фосфатов различными частями корневой системы Hordeum vulgare L. (ячмень).Планта
139
,47
–53.Clarkson DT, Scattergood CB.
1982
. Рост и транспорт фосфатов в растениях ячменя и томата во время развития фосфатного стресса и восстановления после него.Журнал экспериментальной ботаники
33
,865
–875.Кольятти DH, Кларксон DT.
1983
. Физиологические изменения и поглощение фосфата растениями картофеля во время развития и восстановления после дефицита фосфата.Physiologia Plantarum
58
,287
–294.{{-}} \) флюсов вдоль корней риса и кукурузы.Завод, клетки и окружающая среда
21
,240
–246.Дрю М.С., Балобан Л.Р., Барбер С.А., Дженкинс В.
1984
. Изменения кинетики абсорбции фосфатов и калия корнями ячменя с дефицитом питательных веществ, измеренные методом истощения запасов раствора.Планта
160
,490
–499.Dunlop J, Phung HT, Meeking R, Белый DWR.
1997
. Кинетика, связанная с абсорбцией фосфата Arabidopsis и ее регулирование статусом фосфора.Австралийский журнал физиологии растений
24
,623
–629.Эллиот Дж., Линч Дж. П., Лаучли А.
1984
. Приток и отток фосфора в корни интактных растений кукурузы. Двойная маркировка с 32 P и 33 P.Физиология растений
76
,336
–341.Эпштейн Э.
1972
. Минеральное питание растений: принципы и перспективы . Нью-Йорк: Вили.Эпштейн Э., Шмид В.Е., Рейнс Д.В.
1963
. Значение и техника краткосрочных экспериментов по поглощению растворенных веществ тканями растений.Физиология растений и клетки
4
,79
–84.Эшель А., Вайсель Ю.
1996
. Многообразие и многофункциональность различных составляющих одной корневой системы.В: Waisel Y, Eshel A, Kafkafi U, eds. Корни растений, скрытая половина. Нью-Йорк: Марсель Деккер Инк., 175–192.Фергюсон И.Б., Кларксон Д.Т.
1976
. Поглощение ионов в связи с развитием подкожной клетчатки корня.Новый фитолог
77
,11
–14.Слесарь А.
1991
. Экологическое значение архитектуры корневой системы. В: Аткинсон Д., изд. Рост корней растений. экологическая перспектива. Оксфорд: Научные публикации Блэквелла, 229–246.Gao S, Pan WL, Koenig RT.
1998
. Интегрированный возраст корневой системы в зависимости от активности усвоения питательных веществ растениями.Агрономический журнал
90
,505
–510.Хенриксен Г. Х., Блум А. Дж., Спансвик, РМ.
1990
. Измерение чистых потоков аммония и нитратов на поверхности корней ячменя с помощью ионоселективных микроэлектродов.Физиология растений
93
,271
–280.Холобрада М.
1977
. Изменения поглощения и накопления сульфатов вдоль первичного корня во время дифференцировки тканей.Biologia Plantarum
19
,331
–337.Jackson RB, Manwaring JH, Caldwell MM.
1990
. Быстрая физиологическая адаптация корней к локальному обогащению почвы.Природа
344
,58
–60.Джексон В., Квик К.Д., Фольк Р.Дж., Бутц Р.Г.
1976
. Приток и отток нитратов интактными проростками пшеницы: влияние предшествующего нитратного питания.Планта
132
,149
–156.Юнгк А.
1987
. Взаимодействие почвы и корня в ризосфере, влияющее на доступность фосфора для растений.Журнал питания растений
10
,1197
–1204.Юнгк А., Ашер С.Дж., Эдвардс Д.Г., Мейер Д.
1990
.Влияние фосфата на кинетику поглощения фосфата кукурузой ( Zea mays ) и соей ( Glycine max ).Растения и почвы
124
,175
–182.Kochian LV, Shaff JE, Kuhtreiber WM, Jaffe LF, Lucas WJ.
1992
. Использование внеклеточной ионоселективной системы вибрирующих микроэлектродов для количественного определения потоков K + , H + и Ca 2+ в корнях кукурузы и клетках суспензии кукурузы.Планта
188
,601
–610.Краусс М., Фуседер А., Бек Э.
1987
. Развитие и восполнение зоны обеднения фосфора вокруг первичного корня кукурузы в течение вегетационного периода.Растения и почвы
101
,247
–255.Ли РБ, Рэтклифф РГ.
1993
. Внутриклеточное распределение неорганического фосфата и уровни нуклеозидтрифосфата в зрелых корнях кукурузы при низкой внешней концентрации фосфата: измерение с помощью 31 P-ЯМР.Журнал экспериментальной ботаники
44
,587
–598.Ляо Х., Рубио Дж., Ян Х, Цао А, Браун К.М., Линч Дж. П..
2001
. Влияние доступности фосфора на глубину прикорневой части бобов обыкновенной.Растения и почвы
232
,69
–79.Lynch JP.
1995
. Корневая архитектура и продуктивность растений.Физиология растений
95
,7
–13.Линч JP, Браун К.
2001
. Кормление верхнего слоя почвы — архитектурная адаптация растений к низкой доступности фосфора.Растения и почвы
237
,225
–237.Lynch JP, Epstein E, Lauchli A, Weigt GI.
1990
. Автоматизированная система выращивания песка в теплице, подходящая для изучения питания P.Завод, клетки и окружающая среда
13
,547
–554.Lynch JP, Nielsen KL.
1996
. Моделирование архитектуры корневой системы. В: Waisel Y, Eshel A, Kafkafi U, eds. Корни растений, скрытая половина. Нью-Йорк: Марсель Деккер Инк., 247–257.Lynch JP, van Beem JJ.
1993
. Рост и архитектура корней проростков генотипов фасоли обыкновенной.Crop Science
33
,1253
–1257.McNaughton S, Chapin III FS.
1985
. Влияние фосфорного питания и дефолиатона на злаки C 4 с равнин Серенгети.Экология
66
,1617
–1629.Moran CJ, Pierret A, Stevenson AW.
2000
. Поглощение рентгеновских лучей и фазово-контрастное изображение для изучения взаимодействия между корнями растений и структурой почвы.Растения и почвы
223
,101
–117.Muchhal U, Raghothama KG.
1999
. Транскрипционная регуляция переносчика фосфата растений.Proceedings of the National Academy of Sciences, USA
96
,5868
–5872.Ньюман И.А., Кочян Л.В., Грусак М., Лукас В.Дж.
1987
. Потоки H + и K в корнях кукурузы: характеристика и стехиометрия с использованием ионоселективных микроэлектродов.Физиология растений
84
,1177
–1184.Nielsen JS, Joner EJ, Declerck S, Olsson, S, Jakobsen I.
2002
. Фосфовизуализация как инструмент для визуализации и неинвазивного измерения динамики транспорта фосфора в арбускулярных микоризах.Новый фитолог
154
,809
–819.Ниссен П.
1996
. Механизмы поглощения В: Waisel Y, Eshel A, Kafkafi U, eds. Корни растений, скрытая половина. Нью-Йорк: Марсель Деккер Инк., 511–527.Овусу-Бенноа Э, Уайлд А.
1979
. Авторадиография зоны обеднения фосфатом вокруг корней лука при наличии везикулярно-арбускулярной микоризы Glomus mosseae .Новый фитолог
82
,133
–140.Pothuluri JV, Kissel DE, Whitney DA, Thien SJ.
1986
. Поглощение фосфора слоями почвы с разными уровнями фосфора в почвенных тестах.Агрономический журнал
78
,991
–994.Превот П., Стюард ФК.
1936
. Особенности корневой системы в связи с проблемой поглощения солей.Физиология растений
11
,509
–534.Rovira AD, Bowen CD.
1970
. Транслокация и потеря фосфата по корням проростков пшеницы.Планта
93
,15
–25.Rubio G, Liao H, Yan X, Lynch JP.
2003
. Кормление верхнего слоя почвы и его роль в конкурентоспособности растений за фосфор обыкновенной фасоли.Crop Science
43
,598
–607.Rubio G, Oesterheld M, Alvarez CR, Lavado RS.
1997
. Механизмы увеличения поглощения фосфора переувлажненными растениями: доступность фосфора в почве, морфология корней и кинетика поглощения.Oecologia
112
,150
–155.Rubio G, Walk T, Ge Z, Yan X, Liao H, Lynch JP.
2001
. Корневой гравитропизм и подземная конкуренция между соседними растениями: подход к моделированию.Анналы ботаники
88
,929
–940.Рассел Р.С., Кларксон Д.Т.
1975
. Ионный транспорт в корневых системах. В: Сандерленд N, изд. Перспективы экспериментальной биологии , Vol. 2. Оксфорд: Pergamon Press, 401–411.Рассел Р.С., Сандерсон Дж.
1967
. Поглощение питательных веществ разными частями неповрежденных корней растений.Журнал экспериментальной ботаники
18
,491
–508.Сингх С.П., Гептс П., Дебук Д.Г.
1991
.Расы фасоли обыкновенной ( Phaseolus vulgaris, Fabacea).Экономическая ботаника
45
,379
–396.Зильбербуш М.
1996
. Моделирование поглощения ионов почвой. В: Waisel Y, Eshel A, Kafkafi U, eds. Корни растений, скрытая половина. Нью-Йорк: Марсель Деккер Инк., 643–658.Smethurst PJ, Comerford NB.
1993
. Моделирование поглощения питательных веществ отдельными или конкурирующими и контрастирующими корневыми системами.Журнал Американского общества почвоведов
57
,1361
–1367.Snapp SS, Koide R, Lynch JP.
1995
. Использование локализованных участков фосфора корнями фасоли обыкновенной.Растения и почвы
177
,211
–218.Snapp SS, Lynch JP.
1996
. Распределение и ремобилизация фосфора в фасоли под влиянием фосфорного питания.Crop Science
36
,929
–935.Tinker PB, Nye PH.
2000
. Движение растворенных веществ в ризосфере . Нью-Йорк: Издательство Оксфордского университета.Waisel Y, Eshel A.
1992
. Различия в захвате ионов корнями разных типов.Журнал питания растений
15
,945
–958.Wanke M, Ciereszko I, Podbielkowska M, Rychter AM.
1998
. Ответ на дефицит фосфата в фасоли ( Phaseolus vulgaris L.) корни. Респираторный метаболизм, локализация сахара и изменения ультраструктуры клеток корня фасоли.Анналы ботаники
82
,809
–819.Уайт П., Банфилд Дж., Диас М.
1992
. Однонаправленные потоки Ca 2+ в корнях ржи ( Secale cereale L.): сравнение вырезанных корней с корнями интактных растенийJournal of Experimental Botany
43
,1061
–1074.Yeung EC.
1984
. Авторадиография. В кн .: Василь И.К., под ред. Клеточные культуры и генетика соматических клеток растений . Орландо: Academic Press, 778–784.Журнал экспериментальной ботаники , Vol. 55, № 406, © Общество экспериментальной биологии, 2004; все права защищены
Корневое развитие овощных культур: Глава XX
Корневое развитие овощных культур: Глава XX КАТАЛОГ БИБЛИОТЕКИHOME AG СОДЕРЖАНИЕ СЛЕДУЮЩАЯ ГЛАВА
ГЛАВА XX. ФАСОЛЬ
Бобы относятся к семейству растений ( Leguminosae ). который, за исключением, возможно, травы, занимает первое место в сельскохозяйственном значении.Есть много родов, видов и сорта фасоли. Самые важные бобы, выращиваемые в США Состояниями являются фасоль ( Phaseolus vulgaris, ) и лима. фасоль ( Phaseolus limensis и P. lunatus ). В отличие от горох — морозостойкая культура, бобы чувствительны к заморозкам и требуют теплой почвы и теплого времени года для их роста.
ОБЫЧНАЯ ИЛИ ПОЧЕЧНАЯ ФАСОЛЬФасоль — однолетнее высокорослое растение. Карликовые сорта или кустарниковые бобы ( P.vulgaris humilis ) являются низкого роста и непривычки лазать. Это, безусловно, самый важные виды фасоли, выращиваемые в США. Нашлось почти в каждом огороде и широко выращивается на коммерческая основа. В Америке существует 150 разновидностей фасоли. 66
Сорт Wardwell’s Kidney Wax использовался для коренные исследования. Это разновидность кустовой фасоли с восковыми стручками. Урожай был посажен 18 мая, ранее посев замерз. Растения размещали на расстоянии 6 дюймов друг от друга во 2 рядах.На расстоянии 5 футов.
Ранняя разработка. — Саженец фасоли характеризуется очень сильным развитием стержневого корня. Растения только в стадии семядоли, но выращивается в теплой, рыхлой почве, часто имеют хорошо разветвленный стержневой корень длиной 12 дюймов.
Впервые было исследовано развитие корней в поле 18 июня. Растения были уже от 5 до 6 дюймов в высоту и имели разброс 10 дюймов. У каждого было около 10 листов, причем листовки были От 3 до 4 дюймов в длину и от 2 до 3 дюймов в ширину.Общий лист площадь в среднем составляла 1,4 квадратных фута.
Корневая система характеризовалась сильным, стержневой корень почти вертикально нисходящий и множество длинных горизонтальных ветви, возникающие в основном на уровне почвы от 2 до 6 дюймов (Рис. 53). Некоторые из этих больших ветвей, как у лимской фасоли, были придаточными, возникающий из основания стебля. Хотя несколько горизонтальных корни были более 2 футов в длину, их путь был настолько извилистым, что максимальное поперечное распространение было чуть меньше 2 футов.Обычно только около 25 корней на одно растение превышали длину 6 дюймов. Короче их было очень много. Например, между глубинами 3 и 8 дюймов обычно находили в общей сложности 21 корень на дюйм, хотя на большей глубине их было меньше (например, 7 на дюйм между 9 и 14 дюймами включительно).
Рис. 53. Месячная корневая система фасоли Wardwell’s Kidney Wax.
Как правило, многие из более коротких горизонтальных корней, всего от 2 до 4 дюймов в длину, были неразветвленными или плохо меблированы с боковинами.На более длинных ветвление происходило в среднем из расчета 10 веточек на дюйм. Они варьировались от 0,2 до 2 дюймов. в длину. Некоторые были от 4 до 8 дюймов в длину и обладали ветвями. третьего порядка. Таким образом, почва на глубину 8 дюймов была довольно хорошо занят сеткой корешков. Они показаны на рис. 54, который представляет собой вид поверхности корней в первых 6 дюймах почвы.
Рис. 54. — Вид сверху корни фасоли в первых 6 дюймах почвы.Сравнить с Рис.53.
Сравнение горизонтального обзора с вертикальным дает адекватная картина распространения и положения корня. Что корни быстро удлинялись, было видно на 3 дюйма и более неразветвленных концов корня. Тоже будущее, косо нисходящим курсом уже можно было предсказать по положению корневых концов.
Главный корень ниже уровня 8 дюймов был хорошо одет. с корешками (рис. 53). Только те, которые превышают 3–6 дюймов в длина была изменена.Последние 5 дюймов набухшего, толстого, белые стержневые корни были довольно гладкими, хотя и значительно изогнутыми и изогнутые от проникновения в плотную почву. Максимальная глубина 27 дюймов.
Рост в середине лета. — Второй экзамен был произведен примерно через месяц, 14 июля. почти 1 фут в высоту и примерно 1 фут в ширину. Те среднего размера имел около 18 крупных листьев, самый крупный из которые были более 1 фута в ширину. Самые большие листочки имели длину и диаметром 6 и 5 дюймов соответственно.Просвечивающая поверхность увеличился до 5,5 квадратных футов. Растения обильно цвели и бурно росли многочисленные молодые плоды.
Рис. 55. Корневая система почек. фасоль около 2 месяцев.
Корни тоже значительно выросли, хотя это было не так заметно, как у многих других овощных культур. Боковой разброс достигал максимума 30 дюймов, наиболее широко распространенный корни иногда заканчиваются на поверхности почвы.
Где кончики корней были повреждены или разрушены, от концов корней отходили многочисленные боковые части, а некоторые из них следовали курсом, который главный корень. Этот феномен роста боковых стволов продвигается. повреждением основной ветви неоднократно наблюдалось среди много овощных и полевых культур.
Стержневые корни также увеличили свою глубину с около 27 дюймов на июньском осмотре до 3 футов. Точно так же рабочий уровень (без стержневого корня), который едва превышает 12 дюймов, теперь была расширена примерно до 20 дюймов в глубину.
Как и прежде, значительные вариации были обнаружены в общее количество крупных основных корней. На заводе, выбранном для рисунок, большинство из них, как и в случае с фасолью, возникли из основания увеличенного стебля. Иногда стержневой корень давал больше их. Сравнение рис. 53 и 55 показывает, что корни на растения, выбранные для представления более поздней стадии, вряд ли горизонтально, как на предыдущем рисунке. Во многом это связано с к вариациям в лицах. Нисходящий рост этих широко раскидистые, а также более наклонно растущие корни здесь полным ходом.
Эти сильные основные или боковые корни имели диаметр от 2 до 3 миллиметров и толщиной не менее 1 миллиметра на всем протяжении их курс. Скорость ветвления, как и прежде, составляла около 10 боковых сторон. на дюйм корня. Их было от 3 до 15, но обычно они приближались большее число. Многие из них были всего от 0,2 до 1 дюйма в длину, особенно на молодых частях старых корней и на более длинные ветви старых. Последние были теперь в изобилии и варьировались от более 1 дюйма до 6 дюймов в длину.Обычно они были обильно разветвленные, нередки корни четвертого порядка. Часто ветви располагались группами по три на противоположных стороны корней. Общее направление этих крупных ветвей, их рыхлый и часто извилистый ход, а также степень разветвленности лучше всего можно визуализировать, изучив рисунок.
Что касается стержневого корня, он играет важную роль. Заполнение почвы многочисленными ветвями прямо под растением и значительно выходя за рамки рабочего уровня другого корни, она постоянно привлекала новые источники снабжения.Несмотря на то что некоторые стержневые корни были вертикальными в направлении роста, другие следовал очень окольным путем, как показано на рисунке. Как прежде, много коротких ветвей пришло из более старой, более мелкой части. Эти были лучше перегруппированы, чем при более ранней экспертизе. Более того, многочисленные, длинные, хорошо разветвленные боковые стороны, идущие в разные стороны, развилась и значительно увеличила поглощающую площадь. Как и на всех основные корни, а также многие из более мелких, 3 или несколько дюймов безветвленных блестящих белых кончиков показали, что растения все еще быстро росли.
Зрелые растения. — Последнее исследование почек фасоль была выращена 5 августа. Очень лиственные растения были около 13 дюймов. высокий и имел общий разброс 15 дюймов. Хотя некоторые еще были Цветущие, большинство из них имели обилие зрелых стручков. Который они были полностью выросли, что было показано высыханием некоторых листьев.
Корни не увеличились в боковом распространении, но стержневые корни и некоторые из самых длинных основных отводов (включая возникающие случайно из ствола) простирались до глубины от 40 до 46 дюймов, рабочий уровень также был значительно увеличен, я.е., от 20 до 36 дюймов. Разветвление корней было очень хорошо развиты до уровня 3 фута и на расстоянии 2 фута со всех сторон растения. Многие корни уходили глубже. Немного, особенно более глубокая часть стержневого корня, показывала признаки разлагаться.
Сводка. — Фасоль быстро развивается стержневой корень глубоко проникающий. На растениях возрастом 31 день и 6 дюймов высокий, он достигает в длину более 2 футов. Ветвление обильное повсюду, но особенно в 10 дюймах от поверхности.Корни от основания стебля и от стержневого корня проходят горизонтально. но коварно на втором-восьмом дюйме почвы на расстояния от 12 до 24 дюймов. С многочисленными ветвями они составляют большая часть поглощающей системы, которая является отчетливо поверхностной. Через месяц, когда цветение будет обильным и начнутся плоды. для формирования корни более обширные. Главный корень увеличился его глубина до 3 футов и рабочий уровень до 20 дюймов. это широко разветвленный даже на втором футе почвы.Основная горизонталь ветви расширили боковое распространение до 30 дюймов и имеют ветви, глубоко проникающие во второй фут почвы. Когда растения почти созрели, почва до 2 футов со всех сторон хорошо разветвлен до рабочего уровня 3 фута и имеет многочисленные корни вытяните на 1 фут глубже.
Таким образом, общая корневая характеристика фасоли не сильно отличается от гороха, хотя боковое распространение и глубина проникновения несколько больше и чем глубже почва, прямо под растением, более тщательно занятый.
ЛИМА БОБЫЛима большая ( Phaseolus limensis ), хотя это многолетнее растение на юге, где оно значительно коммерческое значение, выращивается как однолетник на севере США. Состояния. Это энергичный производитель, но требует более высоких температур. и более продолжительный вегетационный период, чем у обычных садовых бобов. это крепкое, высоко вьющееся растение, хотя кустарниковые или карликовые формы не взбираться.
Бобы Буша Лима Берпи (сорт limenanus ) крупносемянного плоского типа был посажен 2 июня рядами 30 дюймов. далекий.Семена помещали в ряд на расстоянии от 6 до 12 дюймов.
Ранний рост. — Корневая система была первой осмотрена через 2 недели, 18 июня. Семядоли пожелтели. и почти у каждого растения было по два листа 4 дюйма в длину и половину ширины. Общая поверхность листа составляла 24 квадратных дюйма. Растения в среднем были 5 дюймов в высоту.
Прочный стержневой корень, выходящий из основания ствол и достиг глубины 13 дюймов. Очень многочисленные, в основном горизонтальные ветви были самыми длинными сверху (максимум 9 дюймов) и постепенно становился короче на большей глубине, давая конусообразную форму. по форме к корневой системе в целом (рис.56). Корней не найдено на поверхности 2 дюйма почвы, но многочисленные, большие, придаточные корни отходили от основания стебля и широко распространялись чуть ниже этот уровень почвы. Они быстро росли и последние 3 дюйма были совершенно неразветвленными. В противном случае ветки только первого порядка От 0,3 до 2 дюймов в длину (но в основном короткие) происходили со скоростью около 8 на дюйм. Из более старой части главного корня, , т. Е. первые 5 дюймов, боковые стороны поднимались в большом количестве, часто как до 30 на дюйм.Их длина варьировалась от 4 дюймов. на старшей части стержневого корня до 1 миллиметра на младшей часть. Последние 4 дюйма стержневого корня были неразветвленными.
Рис. 56. 16-дневная корневая система фасоли Burpee’s Bush Lima.
Рост в середине лета. — Примерно через 5 недель 24 июля было произведено повторное обследование. Заводов было 10 дюймов в высоту и цветет. На толстых стеблях было около 33 уходит каждый. Самых крупных листовок было 3.5 на 2 дюйма в длину и шириной соответственно, а общая светящаяся поверхность составляла 4,3 квадратных фута.
Корневая система также хорошо разрослась. В боковое распространение увеличилось до более чем 2 футов, а глубина почти Было достигнуто 3 фута. Корневая система обычно состояла из стержневой корень и от 6 до 8 больших боковых сторон, отходящих от основания стебель увеличенный, хотя всегда имелись многочисленные более мелкие корешки. Главный корень шел несколько хитрым нисходящим курсом, часто изгибаясь и поворачиваясь в длинные изящные изгибы или резко и проник на самую глубину, от 33 до 35 дюймов.Более поверхностный большие корни распространяются в стороны, часто от 1 до 2 футов в поверхности 8 дюймов почвы, а затем повернуть более наклонно или даже вертикально вниз достигал глубин от 20 до 30 дюймов. Еще другие основные корни, более наклонно вниз, заполняя объем почвы между широко распространенные боковые стороны и площадь, занимаемая стержневым корнем и его ветви (рис. 57). Эти большие корни были от 1 до 2 миллиметров. в диаметре и сохраняла эту толщину на протяжении всего пути.
Рис.57. — Корневая система лимы. фасоль в период цветения (24 июля).
Ветвление боковых сторон было несколько изменчивым. но в целом было обильно. Более короткие ветви были простыми, в основном всего от 0,3 до 2 дюймов в длину и обычно скорость от 4 до 12 на дюйм. Более длинные ветви, т. Е. тех длиной более 2–3 дюймов были практически все снабжены с боковыми стенками и со скоростью, аналогичной основной ветви. Часто они были от 9 до 15 дюймов в длину.
Ветвление стержневого корня было очень обильным; 25 к 28 боковых стволов длиной от 0,2 до 4 дюймов проходили на каждом дюйме через первые несколько дюймов своего хода. Ветвление было особенно выражено в срезе борозды (первые 8–9 дюймов, Рис. 57) некоторые из горизонтальные ветви, довольно широко раскидистые и иногда дающие поднимаются к длинным, вертикально проникающим корешкам. На большей глубине они показали отчетливую тенденцию к расширению на несколько дюймов, а затем повернуть вниз, часто параллельно ходу стержневого корня для 1 фут или больше.Самые длинные ветви уже были снабжены третичные корни. Таким образом, растения имели очень эффективное поглощение. система.
Созревающие растения. — Заключительный экзамен был произведен 25 августа. Растения были хорошо развиты и плодоносили. Они были 19 дюймов в высоту, с общим размахом вершин 30 дюймов, и еще не начал сохнуть. Было найдено около 25 веток на растения среднего развития. Они были покрыты изобилием, из больших зеленых листьев. Множество больших стручков длиной почти 5 дюймов и более 1 дюйма шириной, а также множество более мелких. на нормально развитых растениях.Цветение только что прекратилось.
Подземные части претерпели поразительное развитие. Боковое распространение увеличилось вдвое и теперь составляет максимум 4 фута; глубина проникновения увеличена с 3 до 5,5 футов; и рабочий уровень до 45 дюймов. Обычно стебель продлился от 2 до 2,5 дюймов в почву и дал начало глубокому стержневой корень на его конечной части и почти такие же большие боковые стороны рядом с его основанием. Последних обычно было 7 или 8 (иногда их было целых 11), а по длине и степени ветвления они часто были столь же важны, как стержневой корень.В случаях, когда этих боковых корней было меньше (иногда всего 4) стержневой корень дал начало многим сильным, хорошо развитым боковым стволам. В стержневой корень и большие придаточные корни стебля разнообразны толщиной от 1,5 до 3 миллиметров, обычно стержневой корень имеет больший размер. Почти все сохранили этот диаметр впервые. От 12 до 18 дюймов их курса, а некоторые выдерживали его через расстояние до их концов не менее 5 футов. Иногда корень будет сужаться до 1 миллиметра в толщине, а затем снова увеличиваться.
Один стержневой корень обычно преследовал вертикально вниз, хотя и несколько извилистым, курсом и более протяженным глубоко. Большинство других основных корней проходили на глубину от 1 до 3 футов или более. поверхность от 6 до 12 дюймов перед поворотом вниз. Они иногда однако разветвлялись и давали группы от трех до пяти ветвей которые переходили более или менее вертикально вниз в третью или четвертую фут почвы. Более того, почти все эти корни дали начало их горизонтальный ход к вертикально нисходящим крупным ветвям.Некоторые из них выросли всего на 6 дюймов от стержневого корня, другие выросли. более 3 футов от него. Эти корни со своими ветвями полностью занимает почву до рабочего уровня на 45 дюймов (рис. 58).
Рис. 58. — Большая степень Здесь показано созревающее растение лимской фасоли. Двести кубических футов почвы были разветвлены корнями одного растения.
Скорость ветвления была несколько разной, как по количеству и длине боковых сторон, но всегда было обильно.Норма от 6 до 12 на дюйм была обычной, а от 13 до 16 на дюйм — нет. необычный. Меньшие ветви имели длину от 0,1 до 5 дюймов. Большинство из них, особенно более длинные, выше 2,5 футов, с такой же скоростью поставлялись отводы второго порядка. Корни третьего порядка были не редкостью. Более длинные боковые стороны, многие из которых уже упоминались как нисходящие по вертикали, были повторно разветвлены, как и основные корни. Хотя первые несколько дюймов на основных боковых корнях не было так хорошо обставлены ветвями, эта область была очень хорошо занята короткими боковыми стволами (0.5 к 2 дюйма в длину), возникающие из стержневого корня. Например, 72 из них были найдены на глубине от 3 до 9 дюймов. Эти нитевидные наросты обычно были неразветвленными. Между глубинами 8 и 15 дюймов, корни были примерно вдвое меньше, обычно От 1 до 4 дюймов в длину с 10-15 ветвями на дюйм. Это были часто перегруппировывались во второй порядок, таким образом формируя эффективный корневая сеть, окружающая главный корень. Ниже этого на глубину более 2 футов ветвления снова были более многочисленными, и ветви в среднем намного дольше.Они были очень короткими и мало впитывались произошло ниже 4-футового уровня.
Сводка. — Корневая система лимской фасоли очень похож на фасоль. В раннем росте это имеет стержневой корень с очень многочисленными горизонтальными боковыми ветвями; другие возникают случайно из основания стебля. Корень, система конической формы, основание перевернутого конуса начало, примерно на 2 дюйма ниже поверхности почвы. В течение следующих 5 недель около восьми сильных боковых корней, распространяющихся на 2 фута или более в поверхностный слой почвы, обильно ветвящийся.Поворачиваясь вниз, они заканчиваться второй или третьей ногой. Большое количество меньших отводов засыпать почву возле стержневого корня. Главный корень также покрыт длинные боковые стороны почти до конца, примерно на уровне 3 фута, что значительно увеличивает его впитывающую площадь.
К концу цветения корневая система составила дальнейший заметный рост. Широкораспространенные, почти горизонтальные ветви бегите наружу от 2 до 4 футов на поверхности 12 дюймов. Тогда они часто повернуть вниз и, как длинные боковые стороны, возникающие в их горизонтальных конечно, проникают глубоко, часто далеко в четвертую ступню почвы.Более наклонно спускающиеся длинные корни и более короткие ветви возникающие из стержневого корня (длина которого составляла 5,5 футов), заполняют оставшийся объем почвы под растением. Часто главный ветви так же важны, как стержневой корень. Где их мало, более глубокие части стержневого корня лучше разветвляются. Замысловатый сеть мелких корешков встречается на протяжении от 200 до 225 куб. футов почвы, разветвленной одним растением. Боковое распространение корни около 4 футов. Это на 1,5 фута больше, чем у фасоль, а рабочая глубина примерно на 9 дюймов глубже.В большая корневая система лимской фасоли коррелирует с большей развитие вершин.
Другие исследования бобов. — Много исследований были сделаны на основе корней фасоли, особенно в Европе. Некоторые немецкие исследователи обнаружили, что фасоль ( Phaseolus vulgaris) и вика или фасоль ( Vicia faba ) очень похожи по своим коренным привычкам. Оба они образуют четкую определяется стержневой корень, а затем ряды придаточных корней. Последний возникают сначала от основания гипокотиля, но позже, с с возрастом растения они возникают в значительном количестве из более высоких частей основания стебля.В бобе они были обнаружены быть развитым почти так же сильно, как главный корень. Сильнее корневые ветви первого порядка, встречающиеся в большом количестве, особенно в бобах, проникают примерно на ту же глубину, что и основной корень. Глубина проникновения в боб составляла от 40 до 43 дюйма. Боковой разброс варьировался от 20 до 30 дюймов. 89 Аналогичные результаты по глубине проникновения были обнаружены другими следователями. 133
Две разновидности Phaseolus, Pride of Lyon и принцесса Орлеанская, были тщательно изучены в России.Первый имел такую же глубину проникновения корня (25 дюймов) на возраст 45 дней, как и последний, когда он был на 10 дней старше. В начале периода цветения относительная глубина и боковое распространение для двух видов составляло 26 и 10 дюймов для первый и 30 и 12 дюймов для последнего. Дело в том, что разные четко указывалось, что у разновидностей разная степень корнеобразования. У принцессы Орлеанской был более слабый рост корней, который прекратился. развитие раньше, чем более обширная корневая система Гордость Лиона.Окончательная глубина корня составила около 34 и 38 дюймов. а поперечный разброс составлял 12 и 20,5 дюймов соответственно. Начало с периодом цветения горизонтальные корни увеличивались в длину лишь слегка. 123
Боб Boston Dwarf Wax был выращен в Женеве, Н.Ю., а смытые с почвы примерно в середине августа, когда растение приближалось к зрелости. Почва была суглинком, Глубина от 6 до 10 дюймов, подстилаемая прочным гравийно-глинистым грунтом. Более глубокие корни были прослежены до глубины 2 фута и горизонтального одни простирались так же далеко по обе стороны от завода.Алый В то же время был исследован и раннер-боб. Стебли были около 4 футов высотой, и растение было в полном цвету. Тем глубже корни вытянуты до 2,5 футов, а более длинные горизонтальные не менее расстояние 4 фута. Несколько корней выросли в пределах 1 дюйма от поверхности, но подавляющее большинство из них было от 2 до 8 дюймов в глубину. В корневая система была очень похожа на систему восковой фасоли, но явно более обширный. 44
Другие исследования показали, что фасоль, алый бегун и лимонная фасоль — все они характеризуются довольно крепкий стержневой корень.Когда стержневой корень встречает препятствия, он легко сбивается с пути или может распасться на множество ветви. Боковые стороны первого порядка очень многочисленны, развиваются ранний, а топор обильно одет с сильно разветвленными боковинами второго порядка. Боковой разброс оказался больше. чем у гороха. Бобовые растения, которым всего 36 дней, достигли глубины 12 дюймов и поперечное распространение 20 дюймов. 34
Установлено, что корневая система почки bean может быть значительно изменен.В рыхлой, богатой почвы корни могут увеличить глубину своего проникновения за счет способствуя росту стержневого корня и сильного, прикорневого, наружу и корни, проникающие вниз. Это сопровождается уменьшением в отраслевом производстве — прямо пропорционально глубине. Но если при выращивании нарушены кончики корней, боковые исходящие от этих корней, которые уже длинные и толстые, гораздо более ограничен в снижении роста. Таким образом, компонент также имеет способность адаптироваться к мелкой почве.Поскольку более сильные корни препятствуют проникновению глубоко, они производят более многочисленные и более длинные боковые и базальные корни, возникающие позже, преследуют более горизонтальный ход. 86
Бобы фасоли ( Vicia faba ) имеют корневую структуру. очень похож на фасоль темно-синего цвета с сильным, широко раскидистым боковые стороны, отходящие от верхней части стержневого корня и др. косо проникающие ветви из более глубокой части. Это было экспериментально определяется путем выращивания фасоли в ящиках 6 и 14 дюймов.в глубину соответственно, и залили таким же объемом почвы, чтобы стержневые корни были намного короче на мелководье, чем на глубоком грунте, но это число вторичных корней на единицу длины было больше в неглубоких почва. Когда стержневой корень был отрезан, растение не пострадало. при условии, что разрез проходил достаточно глубоко под поверхностью, чтобы стержневого корня оставалось достаточно, чтобы образовалось множество боковых стволов. Обнаружено свободное развитие стержневого корня на значительную глубину. ненужное для наилучшего роста растений в условиях эксперимента.Но где были более глубокие слои почвы для использования стержневой корень необходим, чтобы дать начало более глубокому боковые. Что растение приспосабливается к мелкой почве, изменяя обычная корневая привычка — факт большого значения и значение для сельского хозяйства. 84
В другом эксперименте бобы выращивались в поле суглинка. Когда растения были от 4 до 5 дюймов в высоту, верхние боковые корни были обрезаны близко к растениям на глубину 6 дюймов. Это было сделано в полдень в начале июня.Погода было ясно и жарко после обильных дождей. Вскоре растения увяли и, собственно, увядание было заметно в течение нескольких дней. На некоторых у растений отмерли нижние листья. Растения плохо росли и к 1 июля очень мало увеличился в высоту, когда второй, аналогичный резка была произведена. Они не увяли после второй обрезки, но даже через месяц они были развиты гораздо слабее, чем неповрежденные растения. С другой стороны, обрезка стержневого корня другие растения ниже основных ветвей, i.э., на глубине 6 дюймов, не привело к увяданию, хотя в то время это почва была достаточно сухой. Это продемонстрировали другие такие глубокие эксперименты, обработка почвы дает большую отдачу там, где растения расположены дальше друг от друга, чем там, где они растут близко друг к другу. 85 Связь с повреждением корня очевидна.
Другие исследования фасоли показали, что он был наиболее обильно разветвлен на глубину 4 дюйма, и это самые большие отводы имели длину 10,5 дюймов.Количество боковых стволов первого порядка было около семи на сантиметр стержневой корень в верхней части корневой системы, но в нижней части их было всего около двух на сантиметр. Более глубокая часть корневой системы было только 29 процентов столь же хорошо разветвленной, как более мелкая часть. Таким образом, по сравнению с горохом, исследованным на в то же время максимальная длина ветвей была на 1,1 дюйма меньше, но количество первичных отводов было несколько больше в поверхностный грунт. На первичных и вторичных отводах у гороха ветвей было в два-три раза больше в более глубоких почвы и общей разветвленности на 23% больше (по сравнению с более мелкая часть корневой системы), чем в случае Боб.
Корни больших плоских бобов лимы, выращенных в Калифорния, простирается на глубину от 3,5 до 6 футов. Стержневой корень проникает почти. прямо вниз на эту глубину, вызывая меньшие и более волокнистых корешков через промежутки времени. Где уровень грунтовых вод находится на таком уровне, что корни растений проникают вниз к влаге, которая возникает из-за капиллярности, бобы очень хорошо себя чувствуют и не так сильно страдают от горячего, засушливые дни. Они выдержат очень сухую почву на поверхности после их корни укореняются ниже.В тропическом климате корень иногда бывает большим и мясистым, и растение живет более чем 1 год. 137
Обзор этих исследований по укоренению привычки фасоли ясно указывают на ошибочность нынешней идеи, в литературе по овощеводству обычно выражается, что бобы — культура с мелкой корневой системой. Это правда, что часть корневая система находится прямо под поверхностью почвы, но корни также широко распространяются и глубоко проникают. Среди разновидностей изучаемые существенные различия были обнаружены в укоренении и способность корневой системы приспосабливаться к разным показаны почвенные среды.Дальнейшие исследования этих и другие из очень многочисленных разновидностей прольют много света на поведение корня в различных условиях роста. Как корни откликнуться на их распространение и деятельность в культуре засушливых земель, орошение, различные методы подготовки семенного ложа, глубины и способ внесения навоза и других удобрений, глубины межфазные интервалы и т. д. — это проблемы, требующие дальнейшего изучения. 85 Больше внимания следует уделять их деятельность в недрах. Это будут не только великие научный интерес, но может привести к модификациям и улучшениям современных культурных практик.
Коренная привычка в отношении культурной практики. — широко распространенные корнеплоды бобов тем, что они обильно снабжены бугорками и пластичностью корневой системы адаптируясь к различным условиям, все приводит к выводу что фасоль будет хорошо расти на различных типах почвы. Дело в том, что они хорошо себя чувствуют практически во всех типах легких супесей к тяжелым глинам хорошо известен большинству садоводов. Это общее бобы будут расти на бедных почвах лучше, чем многие другие культуры выражается на просторечии: «Этого достаточно для бобов.» Однако на севере глинистые почвы могут быть слишком влажными и теплыми. медленно, а песчаные почвы могут быть слишком засушливыми.
Растения отвечают развитием корневой системы и урожайностью при выращивании на умеренно плодородных, тщательно подготовленных, хорошо дренированных но влажная и правильно возделанная некислая почва. Бобовое растение особенно чувствителен к плохой обработке почвы и грубому обращению на время выращивания. Хорошее посевное ложе, хорошо измельченное и прочное упакован, позволяет максимально использовать питательные вещества почвы и влажность на протяжении всего вегетационного периода.В плохо подготовленное семенное ложе, волокнистые корни могут погибнуть, и растение будет повреждено частичной или полной утратой этой части корневой системы занимая самую богатую часть почвы. На почвах, которые тоже богатым наблюдается тенденция к слишком большому росту виноградных лоз и, несомненно, компактная корневая система.
Так как фасоль и фасоль являются эпигеальными привычку и крупные семядоли нужно вытащить из земли, тщательная подготовка и хорошая обработка хорошо уплотненного семенного ложа необходимы.Глубокая рыхлая почва способствует более глубокому развитию корней и в то же время более тщательное заселение самых богатых часть субстрата, поверхностный слой почвы. Это до частично для лучшей аэрации и частично для лучшей водоудерживаемости емкость почвы. В засушливом земледелии это раннее перемешивание почвы образует сухой слой почвы. Если это сохраняется, растения будут укореняться только ниже этого слоя. В противном случае многие поверхности корни, несомненно, погибнут в результате поздней засухи.
Под поливом, где необходимо проделать борозду между рядами, чтобы обеспечить возможность полива во время роста урожая, это практика — глубоко возделывать эту часть почвы.Это мера предосторожности, позволяющая сохранить почву сухой и, таким образом, предотвратить корни не распространяются в область, чтобы они не были травмированы когда борозда открыта. 114 Рыхлитель почва вокруг стеблей способствует образованию очень многих побочных корни. Заглубление почвы на глубину от 16 до 18 дюймов рекомендуется для увеличить проникновение влаги в большем количестве и районах. 55 На определенных участках, где фасоль выращивалась для много лет необходимо практиковать севооборот как профилактическое меры против сухой корневой гнили.Это заболевание вызывается почвенным гриб ( Fusarium martii phaseoli ). Он выглядит как красновато-коричневый сухая гниль у основания растения и часто вызывает потерю почти вся часть более тонких корней, а также сморщенные конца стержневого корня.
Раннее выращивание рядом с растением, несомненно, выгодно, но скорость роста боковых корней следует тщательно считается. Приспосабливая свои корни к неглубокой почве, фасоль ограничены в своем ассортименте для обеспечения питательными веществами.Их корни таким образом, неблагоприятное расположение для продолжительной засухи. Растения быстро развиваются и вскоре полностью покрывают землю, предотвращая быстрое испарение влаги из почвы, если она не впитывается сетью поверхностных корешков. Раннее выращивание следует неглубокий, чтобы не повредить корень. Поздний урожай фасоли показал очень небольшое преимущество в выращивании по сравнению с подавлением сорняков соскабливание поверхности. 152
Правильное расположение растений — важная особенность овощной продукции.Скученность снижает рост ботвы и это, в свою очередь, приводит к плохо развитой корневой системе. Больше сорта и растения, выращенные на плодородной почве, где мало опасность засухи или использование орошения более широко разнесены. Если влажность является ограничивающим фактором роста, как в сухое земледелие, широкие междурядья обязательно. Например, в Колорадо при орошении расстояние между рядами составляет 28 дюймов, но на на засушливых землях они расположены на расстоянии от 36 до 42 дюймов друг от друга. Растения на расстоянии от 4 до 6 дюймов в ряду при первом условии и от 10 до 12 дюймов ниже последнего.77
Сверление семян на расстоянии от 4 до 6 дюймов дает лучшее распределение растений, чем размещение пяти-восьми семян на холме от 2,5 до 3 футов. Этот прежний метод хотя бы частично устраняет скученность. Эксперименты показали, что где фасоль были посажены из расчета пять на холм, а холмы 18 дюймов друг от друга в рядах на расстоянии 3 фута, урожай был чуть больше более половины урожая, если бобы были расположены на расстоянии 6 дюймов отдельно в ряду. 28 Садовые бобы обычно расположены слишком близко 38, что может быть легко видно при изучении привычки укоренения.
БИБЛИОТЕКА HOME AG СОДЕРЖАНИЕ СЛЕДУЮЩАЯ ГЛАВА
корней бобов »Все, что вам нужно знать
Глубина корня
Хотя многие люди считают, что бобы имеют неглубокие корни, это не совсем так. Масса корневого комка варьируется в зависимости от разновидности. Стручковая фасоль (зеленая или стручковая) обычно имеет корневой ком, уходящий в землю примерно на 18-24 дюйма. Однако глубокие корни-кормушки могут достигать глубины от 36 до 48 дюймов в почву. Корни бобов Лимы — как кустовые, так и полевые — могут вырастать более чем на 48 дюймов в длину.
Азотная фиксация
Бобы полезны для почвы. Корни бобов имеют по всей длине узелки из ткани. Большинство узелков сосредоточено вокруг главного корня. Почвенные бактерии извлекают азот из воздуха и «скармливают» им корни бобов, которые накапливают азот в клубеньках. Когда вы собираете урожай фасоли, лучше срезать растение на уровне земли. Это оставляет азот в земле.
Корни съедобных бобов
Помимо стручков, некоторые бобы также дают съедобные корнеплоды.К ним относятся:
- Картофельная фасоль или земляной орех — многолетнее растение, используемое коренными американцами аналогично картофелю.
- Бобы стручковые — у этих растений эффектные цветы и съедобные бобы, а также крахмалистые съедобные корни.
- Гиацинтовые бобы — у них очень эффектные фиолетовые, розовые или белые цветы (основная причина их посадки), а также съедобные корни.
Насекомые и корни бобов
Личинки бобовых жуков проникают в корни растений.Однако нематоды с корневыми узелками — это насекомые, которые чаще всего атакуют корни бобов. Эти крошечные червеобразные насекомые живут в почве. Наиболее опасны они на песчаных почвах и могут нанести значительный ущерб. Нематоды также оставляют отверстия в корневой ткани, через которые могут проникнуть болезни. Сопутствующие посадки душистых бархатцев как в прошлом, так и в текущем сезоне могут уменьшить популяции нематод.
Болезни корней фасоли
Корневые гнили и грибок являются наиболее распространенными болезнями корней фасоли.Наиболее распространенными грибами, поражающими корни бобов, являются виды Fusarium, Rhizoctonia или Pythium. Грибки могут сохраняться в почве годами, питаясь разлагающейся растительностью от предыдущих культур. К сожалению, эффективных органических фунгицидов для решения этой проблемы нет. Севооборот — одно из лучших средств предотвращения болезней. Не поливайте слишком много воды, так как это может спровоцировать болезнь.
Гипокотиль и основная корневая система фасоли обыкновенной (Phaseolus …
Контекст 1
… как корневые системы отдельных особей могут перекрываться или нет. Растения предварительно проращивали и рассаду пересаживали в контейнеры (диаметром 7 см, глубиной 20 см). Магнитно-резонансная томография (МРТ) проводилась ежедневно в течение 2 недель с использованием того же магнита и радиочастотной катушки в спектрометре, что и в исследовании, описанном на рис. 6. Пример на корневой системе растений фасоли обыкновенной (Phaseolus vulgaris L cv. Fardenlosa shiny; рис. 6) имеет сверхпроводящий магнит с вертикальным отверстием 4,7 Тл (Тл), который может вместить растения / горшки диаметром до 17 см и длиной 2 м.На рисунке 3 показаны трехмерные изображения всей корневой системы и нижней части гипокотиля …
Context 2
…, пересаженных в контейнеры (диаметр 7 см, глубина 20 см). Магнитно-резонансная томография (МРТ) проводилась ежедневно в течение 2 недель с использованием того же магнита и радиочастотной катушки в спектрометре, что и в исследовании, описанном на рис. 6. Пример на корневой системе растений фасоли обыкновенной (Phaseolus vulgaris L .cv. Fardenlosa shiny; рис. 6) имеет вертикальное отверстие 4.Сверхпроводящий магнит 7 Тл (Тл), в который можно помещать растения / горшки диаметром не более 17 см и длиной 2 м. На рис. 3 показаны трехмерные изображения всей корневой системы и нижней части гипокотиля того же человека в возрасте 14 лет (рис. 6а) и 21 день (рис. 6б). Качество изображения позволяет …
Контекст 3
… фасоли обыкновенной (Phaseolus vulgaris L. cv. Fardenlosa shiny; рис.6) имеет сверхпроводящий магнит с вертикальным отверстием 4,7 Тл (Тл), который может вмещайте растения / горшки диаметром не более 17 см и длиной 2 м.На рис. 3 показаны трехмерные изображения всей корневой системы и нижней части гипокотиля того же человека в возрасте 14 лет (рис. 6а) и 21 день (рис. 6б). Качество изображения позволяет детально охарактеризовать архитектуру корня, например, разделить стержневые и базальные корни и их индивидуальное распределение (рис. 6). По нашим оценкам, в настоящее время можно визуализировать корни диаметром до 300 мм. Через 21 день (рис. 6б) корни имеют тенденцию скапливаться в нижней части …
Контекст 4
… (Phaseolus vulgaris L. cv. Fardenlosa shiny; рис. 6) оснащен сверхпроводящим магнитом с вертикальным отверстием 4,7 Тл (Тл), который может вместить растения / горшки диаметром до 17 см и длиной 2 м. На рис. 3 показаны трехмерные изображения всей корневой системы и нижней части гипокотиля того же человека в возрасте 14 лет (рис. 6а) и 21 день (рис. 6б). Качество изображения позволяет детально охарактеризовать архитектуру корня, например, разделить стержневые и базальные корни и их индивидуальное распределение (рис.6). По нашим оценкам, в настоящее время можно визуализировать корни диаметром до 300 мм. Через 21 день (рис. 6b) корни имеют тенденцию скапливаться в нижней части цилиндра, указывая на …
Context 5
… 17 см диаметром и 2 м длиной. На рис. 3 показаны трехмерные изображения всей корневой системы и нижней части гипокотиля того же человека в возрасте 14 лет (рис. 6а) и 21 день (рис. 6б). Качество изображения позволяет детально охарактеризовать архитектуру корня, например, разделить стержневые и базальные корни и их индивидуальное распределение (рис.6). По нашим оценкам, в настоящее время можно визуализировать корни диаметром до 300 мм. Через 21 день (рис. 6b) корни имеют тенденцию скапливаться в нижней части цилиндра, что указывает на то, что размер контейнера уже может быть ограниченным для относительно молодого растения. Проекция распределения корней в осевой плоскости (рис. 6c, d) дает более простой …
Контекст 6
… гипокотиль того же человека в возрасте 14 лет (рис. 6a) и 21 день (Рис. 6б). Качество изображения позволяет детально охарактеризовать архитектуру корня, например, разделить стержневые и базальные корни и их индивидуальное распределение (рис.6). По нашим оценкам, в настоящее время можно визуализировать корни диаметром до 300 мм. Через 21 день (рис. 6b) корни имеют тенденцию скапливаться в нижней части цилиндра, что указывает на то, что размер контейнера уже может быть ограниченным для относительно молодого растения. Проекция распределения корней в осевой плоскости (рис. 6c, d) обеспечивает более простую визуализацию углового распределения корней и их …
Контекст 7
… 21 день (рис. 6b) корни имеют тенденцию к упаковка в нижней части цилиндра, что указывает на то, что размер контейнера уже может быть ограниченным для относительно молодого растения.Проекция распределения корней в осевой плоскости (рис. 6в, г) упрощает визуализацию углового распределения корней и их развития. …
Руководство по борьбе с вредителями / Общегосударственная программа IPM UC (UC IPM)
Симптомы и признаки
Существует ряд возбудителей корневых гнилей, которые могут вызывать аналогичные повреждения корневых гнилей у растений фасоли.
Черная корневая гниль, вызываемая Thielaviopsis basicola, встречается на черных глазах, а также на других типах сухих бобов.На подземных стеблях и корнях развивается некротическая ткань от коричневого до черного цвета. Трещины часто образуются в некротизированной ткани коры. Грибок можно идентифицировать по наличию темных покоящихся спор (хламидоспор), которые можно увидеть на пораженной ткани с помощью ручной линзы с 20-кратной лупой.
Корневая гниль фузариоза характеризуется поражениями, которые развиваются на подземных стеблях и стержневых корнях. Со временем кирпично-красные поражения становятся коричневыми, и в корковой ткани пораженных участков появляются продольные трещины.Если соскоблить поверхность поражения, в растительной ткани можно увидеть маленькие красные пятна. В тяжелых ситуациях может погибнуть вся корневая система, хотя иногда над поражением могут образоваться новые корни.
Корневая гниль Rhizoctonia иногда возникает на верхних стержневых корнях старых растений в виде дискретных красновато-коричневых поражений. Инфекции растений старше четырех недель встречаются нечасто и обычно вызывают лишь незначительные повреждения.
Корни с сильной корневой гнилью не смогут обеспечить растение достаточным запасом воды в периоды высокого спроса.Слишком много или слишком мало воды может вызвать пожелтение листьев зараженного растения. В общем, зараженные растения имеют тенденцию быть небольшими с пожелтевшими листьями, которые быстро стареют и дают низкие урожаи.
Комментарии к болезни
Корневая гниль фузариоза чаще всего встречается в середине и конце сезона на полях с долгой историей выращивания фасоли. Болезнь мало повреждает нестрессовые растения, но в условиях засухи, плохого питания или кислородного стресса, заболоченной почвы Fusarium solani может быть одной из причин раннего созревания («срезания») и заметного снижения урожайности. .
Все возбудители корневых гнилей могут сохраняться в почве в течение нескольких лет.
Менеджмент
Долгосрочный севооборот с нечувствительными культурами, такими как травы (однодольные), может помочь снизить инокулят в почве. Избегание избыточного полива или длительного стресса от засухи также может помочь справиться с корневой гнилью бобов. Обработка семян возбудителями корневых гнилей Rhizoctonia и Fusarium может помочь справиться с этими заболеваниями. Чтобы предотвратить загнивание корней Thielaviopsis, сажайте в теплой почве.
| Общее название | Количество на акр ** | REI ‡ | PHI ‡ | |
|---|---|---|---|---|
| (пример торгового наименования) | (часы) | (сутки) | ||
| Меры предосторожности в отношении пестицидов Берегите воду Рассчитать летучие органические соединения Защитите пчел | ||||
| Перечислены не все зарегистрированные пестициды. Ниже перечислены пестициды, имеющие наибольшее значение IPM; наиболее эффективные и с наименьшей вероятностью вызывают резистентность находятся в верхней части таблицы.При выборе пестицида учитывайте информацию, касающуюся свойств пестицида и сроков применения, медоносных пчел и воздействия на окружающую среду. Всегда читайте этикетку используемого продукта. | ||||
| Примечание. При использовании обработанных семян обязательно помещайте их в ряд под зернистыми ризобиями. | ||||
| ОБРАБОТКА СЕМЯН | ||||
| А. | ФЛУДИОКСОНИЛ / МЕФЕНОКСАМ | |||
| (Максим XL) | См. Этикетку | 48 * | NA | |
| НАЗВАНИЕ ГРУППЫ РЕЖИМА ДЕЙСТВИЯ (НОМЕР 1 ): Фенилпирролы (12) / Фениламиды (4) | ||||
| КОММЕНТАРИИ: Относится к Rhizoctonia, Fusarium spp. И другим. | ||||
| ** | Смешайте с достаточным количеством воды до полного покрытия. |
| ‡ | Интервал ограниченного входа (REI) — это количество часов (если не указано иное) от обработки до момента, когда в обработанную зону можно будет безопасно войти без защитной одежды. Интервал до сбора урожая (PHI) — это количество дней от обработки до сбора урожая. В некоторых случаях REI превышает PHI. Самый длинный из двух интервалов — это минимальное время, которое должно пройти до сбора урожая. |
| 1 | Номер группыприсваивается Комитетом действий по сопротивлению фунгицидам (FRAC) в соответствии с различными способами действий.Фунгициды с другим номером группы можно чередовать в программе управления резистентностью. Для фунгицидов с номерами групп по способу действия 1, 4, 9, 11 или 17 сделайте не более одного применения перед тем, как перейти к фунгициду с другим номером группы по способу действия; для фунгицидов с другими номерами групп, сделайте не более двух последовательных обработок перед тем, как перейти к фунгицидам с другим номером группы по способу действия. |
| * | REI Исключение: если семена обрабатываются продуктом, а обработанные семена вводятся в почву или засыпаны почвой, Стандарт защиты рабочих при определенных обстоятельствах разрешает рабочим входить в обрабатываемую зону, если не будет контакта с чем-либо, что лечился. |
Руководство UC IPM по борьбе с вредителями: сухие бобы
Публикация UC ANR 3446
C.A. Frate (почетный), UC Cooperative Extension округ Туларе (почетный)
P.G. Gepts, Plant Sciences, Калифорнийский университет в Дэвисе,
R.F. Long, UC Cooperative Extension Округ Йоло
Благодарность за вклад в развитие болезней
A.E. Холл (заслуженный), Ботаника и науки о растениях, Калифорнийский университет в Риверсайде (заслуженный)
R.М. Дэвис (заслуженный), Патология растений, Калифорнийский университет в Дэвисе (заслуженный)
Р.Л. Гилбертсон, Патология растений, Калифорнийский университет в Дэвисе
С. Темпл, Науки о растениях, Калифорнийский университет в Дэвисе,
Текст обновлен: 18.01.
Таблица лечения обновлена: 18.01.
Лопаткомика обыкновенной фасоли — Департамент растениеводства
Шаг 1: Идентификация образца
Отобрать репрезентативные образцы
Шаг 2: выкопайте образец
Это может быть сложнее, чем кажется.Конечно, не стоит обрезать слишком близко к стеблю, потому что тогда у вас не будет достаточно корней, на которые можно было бы смотреть. Но вы также не можете выкопать слишком большую яму, потому что тогда корневой ком сломается и расколется на части, что приведет к срезанию и потере целых частей корней. Все это зависит от текстуры почвы, но мы обнаружили, что около 25 см вокруг стебля, как правило, достаточно большая секция, чтобы хорошо рассмотреть корни, но не настолько большие, чтобы они начали ломаться.
Осторожно окопайте растение или растения, которые вы выкапываете, а затем вставьте лопату под корневой ком и выньте его другой рукой.Затем либо поместите корневой ком в тачку для транспортировки в место мойки, либо поместите его прямо в ванну.
Шаг 3: Замочите, промойте и ополосните образец
Почва, особенно почва с высоким содержанием глины, с трудом удаляется с корней, а поскольку корни бобов особенно хрупкие, необходимо проявлять особую осторожность. Мы наполняем 30-литровые ванны водой с мылом и даем корневому комку постоять не менее 10 минут. Иногда получается образец, который нужно замачивать подольше. По истечении этого периода вы можете осторожно массировать и перемещать распадающийся корневой ком в воде и осторожно смыть всю почву с корней.Часть тонких корней теряется, но лучшего способа избежать этого мы не нашли.
После того, как вы вымыли образец, погрузите его в чистую воду без мыла. Оставьте их в ведре для ополаскивания, пока они не будут готовы к надрезанию, потому что корни бобов быстро высыхают, а затем оставайтесь под тем углом, под которым они оказались во время высыхания. Оставив их в ведре для ополаскивания, корневая система останется максимально увлажненной и неизменной.
Шаг 4. Сделайте снимок образца
.Хотя изображения и близко не так хороши, как на самом деле просмотр образца, и даже несмотря на то, что эти типы изображений бесполезны для компьютерного анализа (см. Метод секционирования корня кукурузы), они все же полезны для грубых сравнений и освежения вашей памяти.Они также помогут вам вспомнить от эксперимента к эксперименту и из года в год, какие баллы вы ставили тем или иным корням.
Мы сделали себе «доску с картинками» из синей разделочной доски и нарисовали на ней черным перманентным маркером. Это помогает лучше выделяться углам, размеру, ветвлению и узловатости, чем на белой доске. Мы нарисовали на доске шкалы для быстрой оценки различных характеристик.
Шаг 5: Оценка образца
Параметры, которые вы оцениваете, как вы их оцениваете и насколько количественными вы хотите быть, полностью на ваше усмотрение.Конечно, лучше всего использовать как можно больше количественных параметров, но чем больше параметров вы действительно измеряете, тем больше времени занимает каждый образец. Если вам нужно просматривать 100 образцов в день, 1 минута более или менее составляет много времени!
Некоторые из признаков, на которые мы обращаем внимание, включают: углы базального корня, базальный диаметр, длину и количество, базальное латеральное ветвление, придаточный диаметр, длину и количество, плотность придаточных ветвлений, диаметр ответвлений, длину и плотность ответвлений, размер узелков, количество хорошие узелки и распределение узелков.Также мы даем оценку болезни (1–9).
Мы попытались найти баланс между количественным измерением и более субъективной оценкой. Например, для оценки длины базальных и придаточных корней интенсивным методом измеряется их диаметр на 1 см от стебля штангенциркулем. Для количества придаточных корней мы подсчитываем количество корней, а для оценки плотности ветвления мы подсчитываем корни в репрезентативном сегменте длиной 2 см. Для ускорения мы просто визуально рассматриваем все параметры, упомянутые выше, и присваиваем им оценку от 1 до 9 (от 1 отлично до 9 очень плохо).Мы также адаптируем наш протокол к типу и размеру эксперимента. Для механистического или физиологического исследования мы проведем более интенсивные количественные измерения. Для большого фенотипического исследования, где мы просто пытаемся увидеть, какие типы корней имеют разные генотипы, мы будем использовать менее интенсивный и менее трудоемкий метод подсчета очков.
Мы сделали себе «доски для подсчета очков», аналогичные «доскам с картинками», но со шкалой (1 хорошо — 9 плохо), точками для размера узелков, которые соответствуют диаметру в миллиметрах (1-5), и с сантиметровыми отметками по краям. для облегчения подсчета плотности ветвления.
Корневые гнили сухих бобов — 2,938
Распечатать этот информационный бюллетень
Х. Ф. Шварца * (5/11)
Краткая информация…
- Распространенными почвенными болезнями сухих бобов в этом регионе являются корневые гнили Fusarium, Fusarium wilt, корневые гнили Rhizoctonia и Pythium damping.
- Корневым гнилям способствуют умеренная или высокая влажность почвы, различные температурные режимы почвы, уплотнение почвы, плохой дренаж, постоянный или частый посев на бобы и другие факторы, вызывающие стресс у растений.
- Патогены годами выживают в инфицированном мусоре и зараженной почве.
- Управление болезнями зависит от севооборота, правильной даты посадки, пестицидов и других рекомендаций
по производству, которые снижают стресс растений.
Многие почвенные грибковые патогены широко распространены в районах выращивания сухой фасоли и фасоли в Колорадо и прилегающих штатах. Потери урожая варьируются от следов до 100 процентов, особенно когда неблагоприятные условия окружающей среды сохраняются после посадки и во время цветения.Наиболее распространенными болезнями и их возбудителями являются гниль корней или сухая гниль фузариоза ( Fusarium solani f. Sp. phaseoli ), фузариозное увядание или желтизна ( Fusarium oxysporum f. Sp. phaseoli ), корень Rhizoctonia и стручок. гнили ( Rhizoctonia solani ) и Pythium, гниение, увядание и стручковая гниль (различные виды Pythium ).
Выживание патогенов
Грибы корневой гнили могут сохраняться в течение многих лет в ранее инфицированных остатках бобов и зараженной почве, создавая перезимовавшие структуры.Это могут быть толстостенные споры ( ооспор Pythium и хламидоспор Fusarium ), гифы (грибковые нити Rhizoctonia ) или небольшие темные склероции ( Rhizoctonia ).
Зимующие структуры стимулируются к прорастанию экссудатом растений из развивающихся чувствительных тканей, таких как корни бобов. Структуры также могут быть стимулированы корнями, не являющимися хозяевами, к безвредному прорастанию или поддержанию
и самовоспроизводству до тех пор, пока чувствительная растительная ткань не станет доступной.Плотность инокулята (количество структур выживания на единицу почвы) этих патогенов может быть уменьшена за счет антагонистических по отношению к ним естественных переносимых через почву организмов и других факторов, описанных в разделе о борьбе с болезнями.
Развитие болезни
Заболеваемость и серьезность каждого грибка корневой гнили и вызываемый ими комплекс болезней зависят от многих факторов окружающей среды, хозяев и культурных факторов. В таблице 1 показаны факторы, которые по отдельности и в совокупности могут способствовать развитию болезни корневой гнили по мере увеличения стресса растений.
| Таблица 1: Описание факторов, способствующих развитию корневой гнили фасоли. | |||
| Фактор | Фузариоз | Ризоктония | Питий |
|---|---|---|---|
| A. Влажность почвы: | по возрастанию | от среднего до высокого | высокая |
| B. Температура почвы: | высокая | низкий — молодые саженцы высокий — в сочетании с холодной поливной водой | от среднего до высокого |
| С.Уплотнение почвы: | твердая плита, плохой дренаж или уплотнение почвы | плохой дренаж | плохой дренаж |
| D. Органическое вещество: | (соотношение углерод / азот) ткань с высоким процентом разложения при посадке | высокая | высокая |
| E. Фертильность: | низкий | неизвестно | неизвестно |
| F. Гербицид: | токсичность, особенно если после посадки прохладно и влажно | токсичность | токсичность |
| г.Вращение фасоли: | менее 3 лет | менее 3 лет | менее 3 лет |
| H. Прочие культуры: | неизвестно | сахарная свекла, картофель | неизвестно |
| I. Плотность растений: | высокая | высокая | высокая |
| J. Качество / возраст семян: | бедные / более 2 лет | бедные / более 2 лет | бедные / более 2 лет |
| К.Выращивание: | обрезка корней | раны корня / гипокотиля | обрезка корней |
| L. Оросительный сток: | внутри / между полями | внутри / между полями | внутри / между полями |
Симптомы болезни
Фузариоз
Рис. 1. Симптомы корневой гнили фузариоза. |
Рисунок 2: Инфекция фузариозного увядания и изменение цвета сосудов. |
Рисунок 3: Пожелтение фузариозного увядания. |
Растения, инфицированные фузариозной корневой гнилью , редко погибают от патогена. Симптомы болезни могут проявляться в секторах круглой или неправильной формы на полях. Растения в этих секторах могут отставать в росте и пожелтеть с ослабленной корневой системой, проявлять преждевременное опадание листьев и плохое заполнение стручков. Однако чаще пораженные растения разбросаны по полю, и наземные симптомы трудно заметить.Первоначальные подземные симптомы проявляются в виде красновато-коричневых полос (поражений) на гипокотиле и первичном корне через две-три недели после появления всходов.
Больные участки растения с возрастом увеличиваются и постепенно буреют. Симптомы могут распространяться вверх по основному корню и гипокотилю на поверхность почвы. В более старых поражениях могут развиться продольные трещины. Сильно инфицированные первичные и вторичные корни обычно погибают, но остаются на растениях в виде высушенных остатков. Боковые корни могут развиваться из гипокотиля над первоначальным местом заражения, если в почве достаточно влаги.Позднее в течение сезона, во время высокотемпературного стресса, сильно инфицированные корневые системы могут проявлять внутреннее изменение цвета (от коричневого до красного) сердцевинной ткани ниже первого узла.
Корневые симптомы Fusarium wilt сходны с симптомами, вызываемыми корневой гнилью Fusarium. Наземные симптомы сначала проявляются на нижних листьях в виде общего пожелтения и увядания, затем становятся более выраженными и переходят вверх к более молодым листьям. Также может наблюдаться задержка роста растений, особенно если заражение растений и высокотемпературный стресс произошли на стадии проростков.Край листа может стать желто-коричневым или коричневым по мере того, как больные растения становятся все более желтыми. Сильно зараженные растения могут демонстрировать стойкое увядание и преждевременное опадание листьев. Наиболее диагностический симптом — это красновато-коричневые или оранжевые полосы в сосудистой ткани до и за пределами первого узла. Обычно это проявляется после появления симптомов на листве во время наполнения стручков в сочетании с высокотемпературным стрессом.
Рисунок 4: Поражения ризоктонией и микросклероции. |
Корневая гниль Rhizoctonia
Корневая гниль Rhizoctonia Симптомы могут проявляться на разбросанных растениях в несколько круглой или неравномерной структуре поля. Грибок может вызвать гибель (отмирание) проростков, корневую и гипокотильную гнили, язвы стеблей и стручковую гниль.
Первоначальные симптомы появляются на корнях или гипокотилях вскоре после посадки в виде линейных или круглых красновато-коричневых затонувших поражений или язв, ограниченных краем от коричневого до красновато-коричневого. Язвы могут увеличиваться с возрастом, становиться более темными и шероховатыми, что замедляет рост растений.Патоген может проникнуть в центральную часть нижнего стебля и вызвать изменение цвета более старых сеянцев до кирпично-красного цвета.
Сильно инфицированные сеянцы или молодые растения могут погибнуть или отломиться на зараженных и ослабленных участках гипокотиля. Поражения также могут развиваться на стручках при контакте с влажной поверхностью почвы и вызывать гниение стручков и обесцвечивание семян.
Питий
Проблемы Pythium обычно разбросаны по полю, поэтому затронутые секторы не образуют узор.Возбудитель может поражать семена, рассаду, молодые и старые растения и стручки. Грибок может вызвать загнивание семян и гибель всходов.
Первоначальные корневые симптомы проявляются в виде удлиненных, пропитанных водой участков на гипокотиле и корнях. Симптомы обычно возникают в течение одной-трех недель после посадки. Вначале инфицированная внешняя ткань стебля становится слизистой и на этом этапе может легко соскользнуть с центральной сердцевины. Однако со временем он высыхает, становится запавшим и приобретает коричневый цвет.
Сильно зараженные растения обычно увядают и умирают.Стручки, контактирующие с влажной почвой, также могут инфицироваться и проявлять водянистую мягкую гниль и массу белого грибкового мицелия (но без образования твердых черных склероций, связанных с заболеванием белой плесенью). Патоген может сильно обрезать корни, снизить общий рост растений и разрушить большую часть гипокотиля и основной корневой системы. Пропитанная водой область на зараженных сеянцах или растениях может выступать на несколько дюймов над линией почвы с небольшими видимыми признаками грибка, если таковые вообще имеются.
Рисунок 5: Корневая гниль Pythium. |
Рисунок 6: Pythium wilt. |
Управление болезнями
С почвенными патогенами сухих бобов и других сельскохозяйственных культур можно бороться, чтобы уменьшить, но не устранить ущерб. Комплексные и тщательно применяемые подходы к растениеводству снижают давление болезней и стресс растений. Это позволяет сильнорослым растениям более успешно получать питательные вещества и влагу в критические вегетативные и репродуктивные периоды.
Севооборот снижает остаточные популяции всех четырех организмов корневой гнили.Лучше всего использовать трехлетний севооборот с посевом бобов каждые три или, в идеале, четвертый или пятый год. Картофель и сахарная свекла могут увеличить количество посевов грибков в почве, и их не следует включать в севооборот, если на поле в анамнезе была корневая гниль Rhizoctonia. Устойчивые культуры включают люцерну, ячмень, пшеницу, овес и кукурузу. Стерня предыдущего урожая всегда должна быть хорошо заделана почвой и разложена перед посадкой фасоли. Это позволяет избежать временного дефицита азота и, следовательно, стресса растений в начале сезона.После тщательного тестирования почвы внесите достаточное количество удобрений, но не в избытке.
Сажайте сухие бобы в теплую влажную почву для быстрого прорастания и появления всходов. Верхний 6-дюймовый профиль должен иметь температуру не менее 60 градусов по Фаренгейту, что чаще всего происходит в этом регионе после 25 мая. Используйте глубину посадки от 1,5 до 2 дюймов, чтобы способствовать быстрому прорастанию. Нормы посадки пегой лошади должны составлять от 60 до 80 фунтов на акр или от 4 до 5 семян на фут, чтобы минимизировать конкуренцию (стресс) между растениями за влажность, питательные вещества и свет без ущерба для урожайности.Всегда применяйте соответствующие гербициды в соответствии с инструкциями на этикетке, чтобы минимизировать повреждение растений, особенно во время прохладной влажной весны.
Посадите высококачественные, обработанные, сертифицированные семена, чтобы максимально увеличить жизнеспособность растений и обеспечить возможность создания насаждений. Большинство сортов в этом регионе одинаково восприимчивы к корневой гнили, но некоторые сорта более устойчивы к заражению фузариозом, чем другие сорта на Западе.
Имеются средства обработки семян и почвы для защиты семян и рассады от инфекции.Внимательно следуйте инструкциям на этикетке. Pythium видов можно обрабатывать с помощью Apron, Ridomil или Maxim. Rhizoctonia solani можно обработать с помощью хлоронеба, топсина, максим и терраклора или PCNB. Первоначальное заражение Fusarium можно отсрочить, но не предотвратить, путем фумигации почвы или обработки Terracoat, Terraclor Super X или Maxim. Эффективность этих пестицидов может быть быстро потеряна, когда условия окружающей среды или культуры становятся слишком неблагоприятными для правильного развития растений.Также ведутся исследования с продуктами биологической борьбы.
Популяции Fusarium и Pythium сокращаются за счет осенней вспашки, а не за счет осеннего или весеннего вскрытия ранее зараженного мусора. Уплотнение почвы, вызванное движением техники и плохим дренажем, может быть уменьшено путем долбления (углубления) на глубину 12-20 дюймов между рядами фасоли или 8-12 дюймов глубиной непосредственно перед сеялкой и на 1,5 дюйма сбоку от ряда. Выполняйте обычные культивации осторожно, чтобы свести к минимуму обрезку корней и, как следствие, стресс растений и увеличение инфекции.Выталкивание почвы вокруг основания растений, инфицированных Fusarium или Rhizoctonia, перед цветением может усилить развитие боковых корней и уменьшить ущерб, нанесенный ранее в результате обрезки корней и инфекции.
Планируйте поливы, чтобы обеспечить достаточным количеством воды развивающиеся (вегетативный рост) и плодоносящие (цветущие и ранние стадии стручков) растения, не вызывая недостатка влаги или избытка влаги. Управляйте поливным стоком, чтобы ограничить распространение корневой гнили и других патогенов внутри полей и между ними. Если стручковая гниль представляет собой проблему, увеличьте интервалы между поливами ближе к концу сезона, чтобы позволить поверхности почвы высохнуть, тем самым подавляя развитие активного грибка и белой плесени.
Следует подчеркнуть, что управление посевами будет наиболее успешным, если производитель объединит рекомендуемые стратегии севооборота, даты посадки, нормы высева, подготовки семенного ложа, сертифицированных семян, обработки фунгицидами, гербицидов и методов выращивания, а также процедур орошения.